扉页
40周年增订版说明
《延伸的表型》(The Extended Phenotype )是我的著作当中唯一一本主要以专业生物学家为写作对象的书,出版于《自私的基因》面世6年之后。对于《延伸的表型》中更富于创新性的章节,我会这样评价:“如果你从没读过我写的任何东西,那就请至少读一读这本书吧。”后来我把这些内容浓缩为《基因的延伸》,作为《自私的基因》第二版的第13章。而《延伸的表型》前面的章节主要是为了从各个不同的角度给后面的内容做好准备和铺垫。其中,第2章和第3章(即本书第14、15两章)特别针对两种对于《自私的基因》的主要误解展开。第2章解决了“伟大的基因决定论谬误”——这是一种错误的解读,认为我们的行为完全是由我们的基因来决定的,而与环境或其他因素无关。第3章解决了“伟大的适应论误解”——所谓“适应论者”所持有的观点,认为生物的所有性状和行为都应被理解为适应性。因为《延伸的表型》这两章完全是由对《自私的基因》的回应而直接产生的,而且它们所强调的一些困惑和误读至今仍存在于某些角落里,于是牛津大学出版社的拉塔·梅农(Latha Menon,我已经与她合作完成了一些卓有成效的工作,出版了好几本书)建议我应该把这两章加入《自私的基因》40周年增订版的最后,形成这本特别的“延伸的”40周年纪念版。我们本来有个好玩的主意,就是想象自己好像面对着一群非生物学专业的听众一样,重新来写作这两章。但是最终,我们觉得还是基本保留这两章最初出版时的原样为好,附带精心挑选过的术语表以助非专业读者理解。这两章展现了人们对于第一版《自私的基因》的一些颇为糟糕的误解,而在接下来的40年间,这两章的讨论应该能够防止类似误解的再次出现。
理查德·道金斯
2016年2月
40周年增订版序言
科学家与政客不同,能够以错为乐。政客如果改变了主张,会被人说成是“反复无常”。托尼·布莱尔(Tony Blair)就夸口说自己“从未开过倒车”。科学家一般来说也愿意看到自己的观点被证明是正确的,但是偶尔一两次的错判同样能够为自己赢得尊重,尤其是当他们能够优雅地承认自己的错误时。我从未听说过有哪位科学家被指责为反复无常。
在某种意义上,我很愿意找到一种方式来收回《自私的基因》一书的核心思想。在基因组学的世界中已经发生了如此之多激动人心的事情,那么一本出版于40年前的以基因为题的书如果还不至于被彻底摒弃的话,必然需要接受大幅度的修改——这似乎是不可避免的事情,甚至很是诱人。然而在这本书中,“基因”的定义比较特殊,它是为进化量身定做的,而不以描述发育问题为目的——若非如此,这本书就真要大改特改了。本书中基因的定义来自种群进化生物学家乔治·C. 威廉斯 [1] ,他已然仙逝,但无疑是本书的英雄。同样离我们而去的还有约翰·梅纳德·史密斯 [2] 和比尔·汉密尔顿。威廉斯认为:“基因是染色体材料上任何一个可能存在得足够长久的代际,并且是可以当作自然选择的单位的部分。”我从这个定义中得出了一个多少有些好笑的结论:“严格来讲,这本书的书名应该是……《染色体有点自私的一大部分以及更为自私的一小部分》。”胚胎学家关心的是基因会如何影响表型,我们新达尔文主义者的关注点则是实体在种群中的频度发生的改变。这些实体在威廉斯看来就是基因(威廉斯后来称之为“抄本”)。基因是可以计数的,而其出现频度是其成功与否的一种测度。本书的一个核心思想是:生物个体不具备上述讨论的基础。单个生物体的基因频度都是100%,因而无法“当作自然选择的单位”。同样,复制单元也无法扮演这样的角色。如果非要说生物个体是自然选择的单位,这其中的意味也很不一样,实际上是把生物个体视为基因的“载具”。生物个体成功与否的测度是其所携带的基因在未来代际中出现的频度,而其奋力去争取最大化的那个量值是汉密尔顿所说的“广义适合度”(inclusive fitness)。
一个基因要获得这些数值层面的成功,就需要在生物个体身上表现出表型效应的价值来。一个成功的基因会在很长一段时期内表现在许多个体身上,它能够帮助这些个体在环境中存活得足够长久,令繁育下一代成为可能。不过,这里的环境指的并不仅仅是身体之外的外部环境,诸如树木、水体、天敌等等,其实还包括了内部环境,特别是其他基因——自私的基因与其他基因共享了一系列生物个体的身体,不仅遍布种群,而且跨越代际。由此,必然的结果是,自然选择会青睐那些在有性繁殖种群中其他基因的陪伴下共同繁荣的基因。从本书的主张来讲,基因的确是“自私的”。但基因同时也能够与其他基因合作,它们所共享的不仅仅只是某一个特定的生物体的身体,还是这个物种的基因库所产生出来的一般意义上的所有身体。一个有性繁殖种群是由相互兼容、彼此合作的基因所组成的联合体——它们今天会合作是由于许多世代以来在先祖类似身体中的合作已经让它们得以繁荣。需要了解的重点在于(这一点常常被人误解),合作性之所以得到青睐,并非是因为一组基因天然地作为一个整体接受选择,而是因为单个基因是单独接受选择的,但这个选择过程的背景是该基因在身体内有可能接触到的其他基因,也就是说物种的基因库内的其他基因。一个有性繁殖物种的每一个个体都是从基因库中抽取自己的基因的。在一系列个体的身体里,这一物种(而非其他物种)的这些基因会不断地彼此相遇,彼此合作。
我们仍不完全清楚究竟是什么原因推动了有性繁殖的起源。但是有性繁殖的一个结果就是,物种可以被创新性地定义为:相互兼容的基因所组成的合作联合体的栖息地。正如在《基因的延伸》这一章所指出的:合作的关键在于,对于每一代而言,一个身体里所有基因共享的那个去往未来的出路如同“瓶颈”一般——那是它们渴望搭乘着前往下一代的精子或卵子。“合作的基因”也是一个同样恰当的书名,而且如此一来,这本书就根本不用做任何修改了,我估计人们因误解提出的许多质疑也可以因此而避免了。
另一个不错的书名是“不朽的基因”。“不朽”比“自私”更有诗意,同时也抓住了本书的一个关键性问题。DNA(脱氧核糖核酸)复制的高保真度对于自然选择而言是一个基本要素,也就是说突变是罕见的。高保真度意味着,基因作为准确的信息拷贝能够存续数百万年之久,成功的基因如此,不成功的基因则不行——而这正是成功性的定义。不过,如果一段遗传信息可能的生命周期很短的话,两者的差异就不会很显著。换个角度来看,每一个活着的生物个体从其胚胎发育期开始就是由一些基因建造出来的,而这些基因能够追溯到许多许多代际以前的许多许多祖先个体身上。活着的生物继承了这些曾经帮助过许多许多祖先存活下来的基因,这就是为什么活着的生物具备存活下去所需的一切,并且能够繁衍下去。它们所需的东西具体是什么,是因物种不同而各异的——捕猎者或是猎物,寄生虫或是宿主,水生或是陆生,栖息在地下或是森林的树冠层——但是普遍性的原则仍是相同的。
本书的一个核心论点是由我的朋友,伟大的比尔·汉密尔顿提出并完善的。我至今仍在为他的离世感到哀痛。我们认为动物应该不仅仅要照料自己的孩子,而且还要照料其他有血缘关系的亲属。对此,有一个我很喜欢的简洁的表述方式,就是“汉密尔顿法则”:如果一个利他性的基因能够在种群中扩散开来,那么必然满足以下条件,即利他者的成本C要小于受益者获得的价值B与两者之间的亲缘度系数r之积。这里的r是一个介于0与1之间的比例。对于同卵双胞胎来说,r的取值是1,子女或同胞兄弟姐妹则是0.5,孙子孙女、异父或异母的兄弟姐妹,以及同胞兄弟姐妹的孩子是0.25,同胞兄弟姐妹生出来的堂或表兄弟姐妹之间则是0.125。但什么情况下r是0呢?在这种定义之下,0的含义是什么?要解释清楚这个问题会更困难一些,但的确是很重要的,而且在《自私的基因》第一版中没有完全给予阐明。0并不意味着两个个体没有共同的基因。所有人类之间共享着超过99%的基因,人与老鼠也有超过90%相同的基因,与鱼有四分之三相同的基因。这些相当高的比例令许多人对于亲属选择产生了误解,其中甚至还包括一些杰出的科学家。但是上面这些数字并非是r的含义。比如说我与我兄弟之间的r值是0.5,那么我与种群背景之中一个可能会与我竞争的任意成员之间的r值就是0。为了对利他主义的进化进行理论化的分析,堂兄弟姐妹之间的r值为0.125只是在与种群参照背景(r = 0)相比较的情况下选取的。这里的参照背景是指种群中其余的个体,利他主义可能也曾在他们身上体现过:虽然面对食物和空间时是竞争对手,但面对物种的生存环境时是历经久远历史的伙伴。这些数值,0.5或是0.125等,指代的是在亲缘关系趋近于零的种群参照背景之上的额外的亲缘关系。
威廉斯所定义的基因是一种可以计数的东西,你可以随着一个物种的代际更替去计算它的数量变化,这无关其分子层面上的本质是什么。比如说,基因的本质是被基本呈惰性的“内含子”(会被翻译机器忽略 [3] )分隔的一系列“外显子”(被表达的部分),然而这一事实并不影响威廉斯所定义的基因。分子基因组学是一个迷人的领域,但是它并未对进化的“基因视角”产生巨大的冲击,而这种视角正是这本书的中心主题。为了说明这一点,我们可以换个说法:《自私的基因》的观点很可能对于其他星球上的生命也是能够成立的,即便那些星球上的基因与DNA毫无关系。不过,现代分子遗传学的具体内容,以及关于DNA的具体研究成果,是可以有办法纳入基因视角之中的。结果你会发现,它们实际上证明了我关于生命的观点的正确性,而非对其产生怀疑。对于这一点,我会稍后再继续探讨。而现在,我要换个跨度有点大的话题,它始于一个非常具体的问题,而这个问题显然能够代表一大票类似的疑问。
你与伊丽莎白二世女王(Queen Elizabeth II)之间的亲缘关系有多近?对我来说,我知道自己是她的十五重堂兄弟姐妹的孙辈。我们共同的祖先是第三代约克公爵理查·金雀花(Richard Plantagenet,1412—1460)。理查的儿子之一是英王爱德华四世(King Edward IV),伊丽莎白女王是他的后代。理查还有一个儿子是克拉伦斯公爵乔治(George,传说是在一桶烈性白葡萄甜酒中淹死的),我是他的后代。很多西方人可能不知道自己与女王的亲缘关系其实比十五重还近,我亦如此,门口的邮递员也是如此。有很多种方法可以让我们成为某个人的远房表亲,或者让我们都成为彼此的亲戚。我知道自己是妻子的十二重堂兄弟姐妹的孙辈(共同祖先是乔治·黑斯廷,第一代亨廷顿伯爵,1488—1544),但是很有可能我们还能通过某种未知的不同方式成为血缘关系更近的亲戚(从各自祖先查下来的不同路径),而且绝对还有许多其他的方式让我们成为血缘关系更远的亲戚。我们所有人都是如此。你和女王可能既是九重堂兄弟姐妹的六世孙辈,又是二十重堂兄弟姐妹的玄孙辈,还是三十重堂兄弟姐妹的八世孙辈。我们所有人,无论生活在世界上的哪个地方,不仅仅是彼此的远房亲戚,而且还有几百种连接亲缘关系的路径。这只不过是用另一种方式表达:我们所有人都是种群的背景,我们之间的亲缘关系指数r趋近于零。我可以依照有据可查的一条路径去计算自己与女王之间的r值,但是根据r值的定义,最终算出来的数值会非常接近于零,因而完全起不了什么作用。
导致所有这些令人头昏脑涨的多重亲缘关系的原因在于性。我们有两位父母,四位祖父母,八位曾祖父母,越往上越多,近乎天文数字。如果你不断乘以2,一直计算到征服者威廉的年代,你的(以及我的、女王的、邮递员的)祖先数量将至少是个十亿数量级的数字,比当时全世界的人口数量还多。这个计算本身就证明,无论你是从哪里来的,我们都共同拥有许多祖先(如果回溯到足够久远的过去,我们的祖先都是完全相同的),所以我们彼此也是很多不同形式的亲戚。
如果你不是从生物个体的视角(生物学家的传统视角),而是从基因的视角(这一整本书都在以不同的方式来提倡这种视角)来看待亲缘关系这件事情的话,所有这些复杂性就都不存在了。不要再问我与我的妻子(或邮递员,或女王)有什么样的亲缘关系,相反,要从单独一个基因的视角来提问,比如我的蓝眼基因:我的蓝眼基因与邮递员的蓝眼基因有什么关系?像ABO血型系统这样的多态性可以回溯到久远的过去,甚至连猿类和猴子也有同样的系统。站在人类A型血的基因的立场上看,黑猩猩身上与之对应的基因是其近亲,而人类的B型血基因反而亲缘关系更远。在Y染色体上的SRY基因决定着动物个体是否是雄性。我的SRY基因会把一只袋鼠的SRY基因“看作”近亲。
或者,我们还可以从线粒体的视角来看待亲缘关系的问题。线粒体是充斥在我们细胞之中的小东西,攸关生死。它们是无性繁殖的,保留了自己残存的基因组(它们与自由生活的细菌有极远的亲缘关系)。根据威廉斯的定义,一个线粒体的基因组可以被看作一个单一的“基因”。我们只能从母亲那里获得线粒体。所以,如果我们想知道自己的线粒体与女王的线粒体有怎样的亲缘关系,那么只会有一个答案。我们可能不知道那个答案是什么,但是我们的确知道她的线粒体与你的线粒体之间成为亲戚的方式只有一种,而非从整个身体的视角来看时的几百种不同方式。你可以一代一代回溯你的祖先,但是只能沿着母亲这条线回溯,你会得到一条单一的(线粒体的)细线,而非不断分支的“整体生物家谱”。你还可以用同样的方法回溯女王的祖先,追踪她的那条母系细线。迟早,这两条线会彼此相遇,此时只要数数每条线上过了多少代,你就能轻易计算出你与女王的线粒体亲缘关系了。
你能在线粒体上做的操作,理论上也可以在任何一个特定的基因上重复,这就显示出了基因视角与生物体视角的不同。从整个生物个体的视角来看,你有两位父母,四位祖父母,八位曾祖父母,等等。但是,每个基因就像线粒体一样,只有一位父母辈,一位祖父母辈,一位曾祖父母辈,等等。我有一个蓝眼基因,女王则有两个。理论上,我们能够回溯基因的代际,找到我的蓝眼基因与女王的每一个蓝眼基因之间的亲缘关系。我们两个基因的共同祖先被称为“聚结点”。聚结分析已经成为遗传学的一个繁盛分支,而且非常吸引人。你看出来了吗?这与整本书都在宣扬的“基因视角”是多么相得益彰啊!我们已经不是在谈论利他主义了。基因视角在其他领域也能显示出自己的力量,比如在寻找祖先这件事情上。
你甚至可以去分析一个生物个体的身体里两个等位基因之间的聚结点。查尔斯王子也有蓝色的眼睛,我们可以假定他的15号染色体上有一对不同的蓝眼等位基因。查尔斯王子的两个蓝眼基因分别来自他的父母,这两个基因之间有多近的亲缘关系呢?对此,我们知道一个可能的答案,而这仅仅是因为王室的家谱比我们大多数人的家谱记录得更为详尽。维多利亚女王也有蓝眼,而查尔斯王子可以有两种方式算作维多利亚女王的后代:从他母亲那边通过国王爱德华七世来算,或从他父亲那边通过黑森的爱丽丝公主来算。有50%的可能性,维多利亚的一个蓝眼基因剥离出了两个拷贝,一个给了她的儿子爱德华七世,另一个给了她的女儿爱丽丝公主。这两个子代基因更进一步的拷贝很容易一代代传到伊丽莎白二世女王和菲利普亲王身上,从而在查尔斯王子身上重新组合在一起。这就意味着,查尔斯两个基因的“聚结点”是维多利亚女王。我们不知道,也不可能知道,查尔斯的两个蓝眼基因是否真是这样的情况。但是从统计学上来说,他的很多对等位基因肯定都会回溯到维多利亚女王身上聚结。同样的推理也适用于你的一对对基因和我的一对对基因。即便我们可能没有查尔斯王子那样清晰记录下来的族谱可供查询,但是你的任何一对等位基因理论上也能找到它们的共同祖先,那就是它们从一个单一先祖基因上“剥离”开的聚结点。
下面要说的是一件有意思的事情。虽然我无法找到自己任何一对特定等位基因的确切聚结点,但是理论上来说,遗传学家们能够对任何一个个体身上的所有对等位基因做聚结点的分析,考虑历史上所有可能的亲缘路径(实际上不是所有的可能路径,因为实在太多了,所以只能是抽取一个统计上的样本),从而得出这个个体整个基因组的聚结模式。位于剑桥的桑格研究所的李恒(Heng Li)和理查德·德宾(Richard Durbin)发现了一件非常棒的事情:一个单一个体的基因组中所有成对等位基因的聚结模式就可以给我们足够多的信息,让我们得以重建其整个物种在之前历史中的种群统计学细节,确定那些重要事件的发生时间。
在我们关于成对基因聚结的讨论中,两个等位基因中的一个来自父亲,一个来自母亲。这里“基因”的含义比分子生物学家所说的基因更为灵活一些。实际上,你可以说聚结遗传学家回归到了一个概念上,有点类似于我之前所说的“染色体有点自私的一大部分以及更为自私的一小部分”。聚结分析研究的对象是一块块的DNA序列,它可能比分子生物学家所理解的单个基因更大,也可能甚至更小,但是它们仍旧能被视为彼此的亲戚,是从一个有限数量的世代之前的共同祖先那里“剥离”出来的。
当一个(上述意义上的)基因“剥离”成为自己的两个拷贝,并分别传给两个子女时,这两个拷贝的后代可能会随着时间的流逝而积累不同的突变。这些突变可能是“不会被注意到的”,因为它们不会表现为表型上的区别。两者之间突变的差异与分离之后经历的时间是成正比的。这一事实,也就是所谓的“分子时钟”,已经被生物学家们很好地利用起来了,可以测量巨大的时间跨度。此外,我们去计算亲缘关系的这些成对基因不一定要有着相同的表型效应。我有一个来自父亲的蓝眼基因,与之成对的等位基因则是来自母亲的棕眼基因。虽然这些基因不同,但它们必定在过去的某个时候有一个聚结点:在那个时刻,我父母的共同祖先的一个特定基因剥离成了两个拷贝,分别传给了这位祖先的两个孩子。这次聚结(不同于维多利亚女王蓝眼基因的两个拷贝)是很久很久以前的事情,从而让这对基因有足够长久的时间来积累差异,尤其是导致不同眼睛颜色的差异。
我刚才提到过,一个生物个体的基因组聚结模式就能够用以重建之前历史中的种群统计学细节。实际上,任何一个个体都可以用来做这件事。碰巧我的整个基因组已经为了录制《性、死亡与生命的意义》这个电视节目进行了测序,这期节目曾于2012年在四频道上播放过。王岩(Yan Wong)是我写作《祖先的故事》(The Ancestor’s Tale )的共同作者,我对于聚结理论的所知以及许多其他方面的知识都是从他那里学到的。王岩利用这个机会对我的基因组做了李/德宾式计算,而且只用了我自己的基因组而已,就推导出了人类的历史。他发现大量的聚结点出现在大概6万年以前。这表明我的祖先所在的那个交配种群在6万年前是一小群人。当时的人口少,那么一对现代基因聚结到当时某位祖先身上的概率就高。较少一些聚结点出现在30万年前,说明当时的有效种群要更大一些。这些数字可以绘成一条曲线,纵轴是有效种群的大小,横轴是时间。下图就是他发现的模式,它与该技术的创始人在任何一个欧洲人的基因组上可以得到的模式都是相同的。
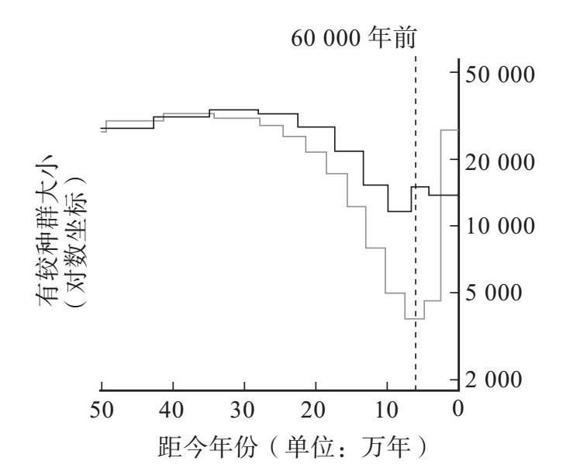
来自理查德·道金斯与王岩合著的《祖先的故事》(2006,第二版)。感谢王岩提供图片。
图中黑线显示了估算出来的不同历史时期的有效种群的大小,依据的是我的基因组数据(我父亲与我母亲之间基因的聚结)。它表明,我的祖先的有效种群大小曾经在大约6万年前显著减小。灰线显示了分析一个尼日利亚人的基因组得到的模式。它显示出差不多同一时期的种群也有所缩小,但程度没有那么剧烈。或许无论是什么灾祸导致了这一下降,它在非洲的下降程度都要弱于在欧亚大陆的下降程度。
顺带一提,王岩曾是我在牛津大学新学院指导的本科生,但后来我从他身上学到的东西多过了他从我身上学到的。本科毕业后,他成为艾伦·格拉芬 [4] 的研究生,而格拉芬也曾是我教过的本科生,后来成为我的研究生,而现在我称他为我知识上的导师。所以,王岩既是我的徒子,又是我的徒孙——这恰恰从文化传承的角度类比了我先前描述的情况,即大家能够以不同的方式成为亲戚。当然了,文化传承的方向要比上述这个简单的构想所代表的情况复杂得多。
总结一下,以基因的视角来看待生命是这本书的一个中心主题,它所阐明的不仅仅是此前版本中详细予以论述的利他和自私的进化,它还阐明了更为久远而深刻的过去,那是我在最初写作《自私的基因》时没能觉察到的。《祖先的故事》第二版中的相关段落(主要是由我的共同作者王岩撰写)为这个问题提供了更为全面的详尽论述。基因视角是如此的强大,以至于一个单一个体的基因组就足以进行详细的定量推算,得出种群的历史性统计数据。它还有什么其他的强大功能吗?正如与尼日利亚人的比较所预示的,未来对于世界不同地域的人所做的此类分析,能够为这种来自过去的种群统计学信号赋予一个地理维度。
基因视角还可能以其他方式穿透历史的迷雾吗?我在几本书中曾经提出了一个想法,我称之为“死亡的基因之书”。一个物种的基因库就是相互支持的基因的联合体,它们曾经一起在过去的特定环境中存活了下来,有久远的过去,也有近世的牵绊,这就使得它们成为那些环境的一种相反印记。如果一个遗传学家具备足够的知识,就应该能够从一种动物的基因组中读出该物种曾经生活的这些环境的特征。理论上来讲,欧洲鼹鼠(Talpa europaea )的DNA应该能够生动地展示出地下世界的面貌,那是一个位于地表之下的潮湿、黑暗的环境,空气中满是蠕虫、腐叶、甲虫幼虫的气味。如果我们知道如何去阅读的话,单峰骆驼(Camelus dromedarius )的DNA将会展现出用编码描绘的古老沙漠,那里有着沙尘暴、沙丘,以及生命对水的渴望。常见的宽吻海豚(Tursiops truncatus )的DNA表达着“深海水域”“快速追逐鱼类”“躲避虎鲸”等信息,不过是以一种我们在未来的某一天可能予以解读的语言写就的。但是同样的海豚DNA中也包含了一些描述其更早时期所处世界的段落,相应的基因也存续了下来:那是它的祖先们在陆地上生活的时期,要小心躲避暴龙和异特龙,活得足够久,久到能够生育才行。在那之前,肯定还有一部分DNA描述了更为古老的生存技能,又一次带我们回到了海洋中,那是它的先祖们还是鱼的时期,被鲨鱼甚至是广翅鲎(巨大的海蝎子)追逐的时代。对于“死亡的基因之书”的研究工作应该会在未来某个时间成为活跃的热点。它会不会成为《自私的基因》50周年纪念版的后记中的精彩内容呢?
[1] 指乔治·克里斯托弗·威廉斯(George Christopher Williams),美国进化生物学家。——译者注(下文如无特殊说明,均为译者注)
[2] 约翰·梅纳德·史密斯(John Maynard Smith),英国著名进化生物学家、遗传学家。
[3] 原文如此。但实际上,内含子与外显子一起被转录成为信使RNA之后,还要经过细胞质内的剪切体的加工,去除不表达的内含子对应区段,才能获得成熟的信使RNA,进入核糖体进行表达。核糖体这个“翻译机器”在此过程中并不会“忽略”内含子。
[4] 艾伦·格拉芬(Alan Grafen),英国动物行为学家和进化生物学家。
30周年版简介
意识到我已经与《自私的基因》一同走过生命中几乎一半的旅程,这把我彻底吓清醒过来,真不知这是件好事还是坏事。这么多年了,我又出版了七本书。每一本书问世时,出版社总派我四处做宣传。读者们以令人受宠若惊的热情回应我的每一本书。他们礼貌地鼓掌,提出一些有智慧的问题,然后排队购书。但他们让我签名的书却是《自私的基因》。这也许有点儿夸张了,有一些读者还是买新书的。而我妻子安慰我说,那些人只是刚刚发现一个新作者,他们会很自然地会去寻找作者的第一本书,阅读《自私的基因》,当然,之后他们肯定会一直读到最新一本书,那才是作者最喜欢的“孩子”。
如果让我宣布《自私的基因》已经严重过时,早已被时代远远抛下,也许我对这种现象会更为耿耿于怀。不幸的是(在某些人的眼里),我没法这么说。虽然有一些细节已经改变,新的例子正层出不穷,但除了一个问题(我马上会来讨论)外,这本书里几乎没有任何内容存在问题,需要我现在急着将它收回,或者需要向读者致歉。利物浦大学的动物学教授阿瑟·凯恩 [1] 是我20世纪60年代在牛津上学时的导师,曾给予我许多启发。他将1976年的《自私的基因》形容为“年轻人的书”。这句话是他故意引用书评家对艾尔(Ayer,英国哲学家)的书《语言、真理与逻辑》的评价。当时的我因此而沾沾自喜,尽管我知道艾尔曾经为其第一本书中大部分内容而公开认错,也不能无视导师的暗示:一段时间后,我也将做同样的事情。
让我从重新思考这个标题开始吧。1975年时,经过我的朋友德斯蒙德·莫里斯(Desmond Morris)的帮助,我将完稿的部分章节交给伦敦出版界的老前辈汤姆·马希勒(Tom Maschler)。我们在乔纳森·凯普(Jonathan Cape)出版社中他的房间里讨论。他表示喜欢这本书,但不喜欢标题。“自私,”他说,“是一个消极的单词。为什么不叫它‘不朽的基因’呢?不朽是一个积极的词,基因信息的不朽是这本书的主题思想,而‘不朽的基因’与‘自私的基因’听起来几乎一样耐人寻味。”(我现在觉得,我们俩都没意识到《自私的基因》刚好呼应了王尔德的《自私的巨人》。)但我现在觉得马希勒也许是对的。许多批评家——特别是那些哗众取宠的批评家(我发现他们一般都有哲学背景)——喜欢不读书而只读标题。也许这个方法足以适用于《兔子本杰明的故事》或者《罗马帝国兴衰史》,但我可以不假思索地说,“自私的基因”标题本身,如果不包含书上大字的脚注,会使人对内容产生一种不恰当的印象。如今,有一个美国出版社无论如何都坚持要求加一个副标题。
解释这个标题最好的方法是标记重点。如果重点在“自私”,你便会以为这本书在讨论人的私心,但是本书却将更多的重心放在讨论利他主义上。这个标题里需要着重强调的词应该是“基因”。让我来解释一下原因。达尔文主义中一直有一个中心辩论议题:自然选择的单位究竟是什么?自然选择的结果究竟是哪一种实体的生存或者灭亡?这个选择的单位多少会变得“自私”。利他主义则在另一个层次才被看重。自然选择是否在种群中选择?如果是这样的话,我们应该能看到个体生物因为“种群的利益”而表现出利他行为。它们将降低生育率以控制种群数量,或者限制其捕猎行为以保持未来种群的猎物储备。正是这个广泛流传的对达尔文主义的误解,给了我写作本书的最初动机。
那么,自然选择是否像我在这里强调的那样,在基因间进行选择呢?在这种情况下,我们便不会惊讶于个体生物“为了基因的利益”,表现出诸如喂养与保护亲属等利他行为,因为亲属更有可能与其共享相同的基因。这种亲属利他行为只是基因自私性在个体利他主义上的一种表现形式。这本书解释了亲属利他与回报——达尔文主义理论中另一个利他行为的主要来源——是如何进行的。如果我要重写这本书,作为一个不久前刚投奔扎哈维/格拉芬(Zahavi / Grafen)“不利条件原理”的人,我会给扎哈维的理论多留点儿位置。扎哈维的想法是:利他主义的捐赠也许是一个“炫富”式的显性信号——看我比你优越好多,我都能负担起给你的捐赠!
让我来重复并扩展一下对书名中“自私”一词的解释。这里的关键问题是:生命中哪一层次是自然选择的单位,有着不可避免的“自私”属性?自私的种属?自私的群体?自私的生物体?自私的生态系统?我们可以争论这些层次中大多数单位的自私性,它们还都曾被一些作者全盘肯定为自然选择的单位。但这都是错误的。如果一定要把达尔文主义简单概括为“自私的某物”,这本书以令人信服的理由层层推理得出,这个“某物”只能是基因。这是我对标题的解释,无论你是否愿意相信推理本身。
我希望这可以澄清那些更严重的误解。尽管如此,我自己也在同样的地方发现了自己犯过的错误。这从第1章中的一句话可以看出来:“我们可以尝试传授慷慨和利他,因为我们生而自私。”传授慷慨与利他并没有错误,但“生而自私”则可能产生误解。我直到1978年才开始想清楚“载体”(一般是生物体)和其中的复制因子(实际上便是基因,第二版中新加入的第13章解释了这个问题)之间的区别。请你在脑海里删除类似这句话的错误句子,并在字里行间补充正确的含义。
这种错误的危险性不难使我认清这个标题的迷惑性,我当时便应该选择“不朽的基因”作为标题。也许这个标题有点儿过于神秘,但所有关于基因与生物体作为自然选择的竞争单位的争议[这个争议一直困扰着晚年的厄恩斯特·迈尔(Ernst Mayr),直至他去世]可以迎刃而解。自然选择有两种单位,它们之间没有任何争议。基因是复制因子的单位,而生物体是载体的单位。它们同等重要,不可低估任何一方。它们代表了两种不同的单位,只有我们认清其区别,才不至于陷入混乱的绝望中。
“合作的基因”是《自私的基因》另一个好的替代书名。虽然这听起来自相矛盾,但这本书主要的一部分便是讨论自私基因的合作形式。需要强调的是,基因组们并不需要以牺牲同伴或者他人的代价来换取自身的繁荣发展。相反,每一个基因在基因库里——生物体以性繁殖洗牌获得的基因组合,以其他基因为背景,追求着自身利益。其他基因是每一个基因生存大环境中的一部分,正如天气、捕食者与猎物、植被与土壤细菌都是环境的一部分。从每个基因的角度上看,背景基因可以与之共享生物体,相伴走过世代旅程。短期看,背景基因指的是基因组中的其他基因。但从长期看,背景基因则是种群基因库内的其他基因。因此,自然选择将基因视作相互兼容——几乎等同于合作——的团体,自然选择偏爱那些共同存在的基因。然而,无论在什么时候,这种合作基因的演化违反了自私基因的根本原则。第5章以桨手的比喻来讲述这个理论,第13章则更进一步讨论了这个问题。
虽然自私基因的自然选择偏爱基因间的合作,我们也必须承认,有一些基因并不这么做。相反,它们牺牲基因组中其他基因的利益。一些作者将它们称为“越轨基因”,有一些则将其称为“极度自私基因”,还有些人直接称之为“自私的基因”,将之与其他因自身利益而合作的基因混为一谈,未能理解其中微妙的不同。第13章讲述了超级自私基因的例子——减数分裂驱动基因,而“寄生DNA”的概念最初是在第3章末尾中提出的,后来有一些作者对其进一步研究,并以“自私的DNA”这样的妙笔来描述。自本书第一次出版,这些年不断有新发现超级自私基因的例子,它们更为匪夷所思。这已成为这些年研究的热点。 [2]
《自私的基因》一直因为将基因拟人化而受到批评,这一点也需要解释一下(如果不需要道歉的话)。我采用了两个层次的拟人:基因与生物体。基因的拟人真不应该是个问题,因为任何有头脑的人都不会认为DNA分子会有一个有意识的人格,任何理智的读者也不会将这种妄想归罪于作者的写作方式。有一次我听到伟大的分子生物学家雅克·莫诺(Jacques Monod)讲述科学中的创造力时,着实心动。我已经忘记了他的用词,但他大概的说法是:当他考虑一个化学问题时,他会问自己,如果我是个电子,我会怎么做?彼得·阿特金斯(Peter Atkins)在其优秀的著作《重临创世》(Creation Revisited )中,在探讨光束通过高折射率介质速度减慢后的折射时,也采取了一个类似的拟人:光束好像想要将其到达终点的时间最小化。在阿特金斯的想象中,这如同海滩边的救生员冲过去拯救一个落水者一样。他是否需要按直线靠近落水者?不需要,因为他跑步比游泳速度更快,在行程中增加陆地行走的比例会更为明智。他是否应该直接跑到海滩边正对着目标的点,来最小化其游泳时间?这个想法好一些,但依然不是最佳方案。通过计算(如果救生员有时间来做这个事情),我们可以找到救生员的最佳行进角度、奔跑距离和不可避免的游泳距离间的最佳组合。阿特金斯总结道:
这正是光线通过密度较大介质时的行为。但光线怎么能在进入之前就已经知道哪一个是最短的行程?它又为什么要在乎这个?
他受量子理论的启发,对这些问题给出了一个绝佳的解释。
这类拟人化的比喻并不只是一种有趣的叙述方式,它还可以帮助职业科学家在雾里看花中,排除错误,找到正确的答案。达尔文主义在利他主义和自私、合作与报复上的计算便是这么一个例子,科学家们很容易推算出错误的答案。但我们经常在最后发现,适当地、小心谨慎地将基因拟人化处理,是将达尔文理论学者从泥沼中拯救出来的最短路径。在本书四大英雄之一的汉密尔顿先驱经验的鼓励下,我自己也尝试着如此谨慎处理拟人化。汉密尔顿在1972年(也是我开始写作《自私的基因》的那一年)的论文里写道:
如果一个基因可以使其复制品聚集起来,形成基因库中一个不断增加的部分,它便会得到自然选择的青睐。我们关注的那些基因会对其携带者的社会行为产生影响。为了让我们的论证更加生动有趣,让我们先试着暂时赋予这些基因以智慧和自由选择的意志。想象一下,一个基因正在考虑问题:如何增加其拷贝。再想象一下它可以有所选择……
这正是阅读《自私的基因》中大部分章节时应有的正确精神。
将生物体拟人化则更加麻烦。这是因为生物体不同于基因,它们拥有大脑,因此也可能真正拥有自私与利他之类主观意识的想法,让我们可以辨识出来。如果本书叫作“自私的狮子”可能会真的迷惑读者,而“自私的基因”不应有这种问题。就像有人可以把自己想象为光束,聪明地选择通过级联透镜与棱镜的最佳路径,或者将自己想象为基因,选择传递千秋万代的最佳路径,我们也可以假定一只狮子计算着其基因长期生存的最佳行为策略。汉密尔顿带给生物学的第一份礼物是其准确的数学计算,这可以算出一只真正的达尔文主义的生物——比如狮子——决定将其基因长期生存的概率最大化时,所应采取的策略。在本书中,我采用了生物体和基因的两个层次,用非正式、口语化的语言来描述这种计算。
在第150页里,我们迅速从一个层次转向另一个层次:
我们已经考虑过在什么条件下母亲让小个子死掉事实上是合算的。如果单凭直觉判断,我们大概总是认为小个子本身是会挣扎到最后一刻的,但这种推断在理论上未必能站得住脚。一旦小个子瘦弱得使其预期寿命缩短到它从同样数量的亲代投资中获得的利益还不到其他幼儿的一半时,它就该体面而心甘情愿地死去。这样,它的基因反而能够获益。
这是个体层次的自我审视。这里的假设不是小个子做出让自己快乐和感觉良好的选择,而是达尔文世界的个体生物会做出的“如果……那么……”的估算,以得出对其基因最有利的选择。这个段落还在继续明确地迅速转化至基因层面的拟人化:
就是说,一个基因发出了这样的指令:“喂,如果你个子比你的骨肉兄弟瘦小得多的话,那你不必死挨活撑,干脆死了吧!”这个基因在基因库中将取得成功,因为它在小个子体内活下去的机会本来就很小,而它却有50%的机会存在于得救的每个兄弟姐妹体内。
接下来的段落则又迅速回到小个子的自我审视:
小个子的生命航程中有一个有去无回的临界点。在达到这一临界点之前,它应当争取活下去,但到了临界点之后,它应停止挣扎,宁可让自己被骨肉兄弟或父母吃掉。
我真的相信,只要读者仔细完整地阅读本书,这两个层次的拟人化一点儿都不会使人迷惑。只要描述恰当,这两个层次的“如果……那么……”评估都会得到完全相同的结论,这也正是判断其正确性的标准。所以,如果我现在重写这本书,我不觉得我会放弃拟人化描述。
重写一遍书是一回事,重读一遍书则是另一回事。我们该如何对待这位澳大利亚读者的判决书呢?
(这本书)非常引人入胜,但有时我希望我没有读过它……一方面,我惊叹于道金斯极为清晰且有根据地看清如此复杂的过程……但同时,我还要责怪《自私的基因》使我在之后的10多年里,不得不与抑郁症进行长期较量……我不再对生命灵魂的认识感到确定,并尝试寻找更深层次的东西——试着去相信,但却不能相信——我发现这本书在字里行间将我所有模糊的想法都一扫而光,而且阻止这些想法重新凝聚于我的脑海中。几年前,这造成了我个人生活中的一次严重危机。
我之前也描述过一些读者产生的类似反应:
我第一本书的一个外国出版商坦言:阅读这本书后,他失眠了3天,被书中传达的冷酷无情的信息深深困扰。另外一些人则问我每天早上如何能离开床铺。一个偏远乡村的教师写信责备我,因为一个学生读完书后含泪找到他,说这本书使她的生命变得空虚而无意义。他建议她不要把这本书给她的任何朋友看,因为他害怕这本书会使他们产生相同的虚无主义思想与悲观情绪。(摘自《解析彩虹》)
如果这些故事是真的,任何良好愿望都无法将其掩盖。这是我要说的第一件事,但我要说的第二件事也一样重要。我在书里接着写道:
想必宇宙的最终命运确实没有意义,但无论如何,我们真有必要将我们生命的希望寄托在宇宙的最终命运上吗?当然不需要,只要我们足够明智。我们的生命被其他更密切、更温暖的人类理想与感觉控制。指责科学剥夺了生命中赖以生存的温暖,是多么荒谬的错误啊,这与我本人及其他科学家的感觉截然相反。我几乎都要对这些大错特错的怀疑绝望了。
另一些批评家则表现出类似“因坏消息到来而迁怒信使”的趋势,他们从《自私的基因》中看到不合心意的社会、政治或经济上的推论,因此反对此书。在1979年撒切尔夫人刚获得其第一次选举胜利后不久,我的朋友史蒂文·罗斯(Steven Rose)在给《新科学家》的文章中写道:
我不是说上奇公司(Saatchi & Saatchi)曾组织一批社会生物学家 [3] 来撰写撒切尔夫人的演讲稿,更不是指一些牛津与苏塞克斯的君子已经开始庆幸终于可以从实际情况解读自私基因这等简单事实,尽管他们一直拼命想要这么告诉我们。这个流行理论与政治事件的巧合要更乱七八糟得多。不过我相信,20世纪70年代末期此书写成时,历史潮流转向了右翼,从法律与秩序转向货币主义与(更为矛盾的)对中央集权的抨击。之后这个转向才成为科学潮流,如果进化理论从类群选择转向亲属选择也能算的话。这个科学潮流变换将被看作推动撒切尔夫人派与其僵化的、19世纪时竞争与排外的人性概念执掌大权的社会潮流的一部分。
“苏塞克斯的君子”指的是不久前去世的约翰·梅纳德·史密斯,史蒂文·罗斯和我都同样欣赏他。史密斯在回复《新科学家》的信中以其典型口吻说:“我们还能怎么做?篡改公式吗?”《自私的基因》传递的一个重要消息(史密斯的文章标题“魔鬼的牧师”更强调了这一信息)是:我们不能把我们的价值观从达尔文主义中推导而来,除非它带着一个消极的信号。我们的大脑已经进化到一个程度,使我们得以背叛自身的自私基因。这种行为的一个明显现象便是我们使用的避孕方式。同样的原理可以也应该作用于更广的范围。
与1989年的第二版不同,30周年纪念版只增加了这篇简介,以及由编辑了我三本书的编辑兼支持者拉塔·梅农选取的一些书评片段,此外并没有新的内容了。除了拉塔外,没有人可以与“K选择”(生态学术语,拥有在环境中获得竞争胜利的能力)超级编辑迈克尔·罗杰斯(Michael Rodgers)媲美。他对此书坚定的信念就像火箭助推器一般,使本书的第一版进入了轨道。
现在这个版本重新采用了最初由罗伯特·特里弗斯 [4] 写作的序言,这也是让我特别高兴的原因。我提过汉密尔顿是本书的四大智囊英雄之一,特里弗斯是另外一个。他的思想贯穿了第9、10、12章的大部分内容,还有第8章的所有内容。他不止给了本书一篇精雕细琢的序言,更不同寻常的是,他选择了本书向世界宣告他超群的新思想:自我欺骗进化的理论。这次他同意让我在此版本中使用原先的序言,我实在感激不尽。
理查德·道金斯
牛津,2005年10月
[1] 指阿瑟·詹姆斯·凯恩(Arthur James Cain),英国进化生物学家、生态学家。
[2] Austin Burt and Robert Trivers(2006),Genes in Conflict: the biology of selfish genetic elemen(哈佛大学出版社),这本书出版时为时已晚,未能列入这个版本的第一次印刷,无疑它是这个重要主题权威的参考书。——作者注
[3] 社会生物学研究生物的社会性行为,通过进化的选择力量来解释生物社会性行为的合理性。这一学科主要聚焦于社会性昆虫的行为研究,但是如果应用到人类社会上就引发了广泛的争议和讨论。爱德华·威尔逊是最早建立这一学科的人,也提出将其应用在人类社会研究中的可能性,前文中提到的史蒂芬·古尔德则对将这一理论应用于人类持批判态度,担心它可能被误解成为基因决定论。
[4] 罗伯特·特里弗斯(Robert Trivers),美国进化生物学家、社会生物学家。
第2版前言
《自私的基因》出版十几年来,书中的主要信息已经成为教科书的正统内容。这其实很矛盾,虽然看起来并不明显,但它并不是那一类作品:出版时因其革命性颠覆而备受指责,而后逐渐稳定获得皈依者,最后被认为无比正统,使人不解最初争议从何而来。《自私的基因》恰与之相反。一开始它得到好评无数,并不被视为富有争议的书。直到数年后,它的争议才逐渐形成。而现在,它被广泛认为是极端的激进作品。但同样在这些年里,当此书极端主义的名声逐渐升级时,它实际的内容则显得越来越不极端,越来越接近通用常识。
自私基因的理论也是达尔文的理论,只是以一种达尔文并未选择的方式来表述。而我也愿意认为,达尔文如果九泉之下有知,也会立刻认识到这种方式的合适性,并为此高兴。这事实上是正统的新达尔文主义的一种逻辑推论,仅仅是以一个新形象展现出来。它并不关注个体生物,而是从基因的视角看待自然界。这是一种不同的观察方式,而不是一种不同的理论。在《延伸的表型》的开篇,我曾用内克尔立方体的比喻来解释这一点。
这是一个二维的纸上墨印图案,但它在观察者眼中却是一个透明的三维立方体。盯着它看上几秒钟,它会变为朝向另一个方向。继续盯着它看,它则会变成原来的立方体。这两个立方体都与视网膜中的那个二维图形同等兼容,于是大脑很乐意在两者间轮流更换。任何一个图形都不比另一个更为正确。我所要说的,便是自然选择有两种观察的方式,可以从基因的角度,也可以从个体生物的角度来观察。如果你恰当地理解两者,它们便是等同的,是同一真理的两种看法。你可以从一者转化到另一者,它依然是相同的新达尔文主义。
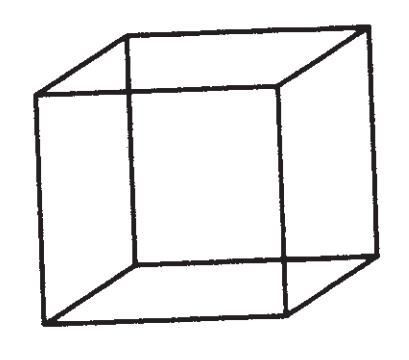
我现在觉得这个比喻太过于小心翼翼了。一个科学家最重要的贡献通常并不是提出一个新理论,或是揭示一个新现象,而是于旧理论和旧现象中发现观察的新方法。内克尔立方体的比喻有误导性,因为它表示两种观察方法的好处是相同的。确切地说,这个比喻还是部分正确的:“角度”和理论不一样,不可以通过实验去验证。我们无法采用熟悉的证明或证伪准则。但在最理想的情况下,视角的改变可以达到比一个理论更崇高的地位。它可以引领整个思想潮流,促使许多激动人心与可验证的理论产生,使之前无法想象的事实暴露无遗。内克尔立方体的比喻完全忽略了这点,它只抓住了视角上改变这一点,却无法公正评价其价值。我们要谈的并不是一个视角的转变,而是在极端条件下的彻底变身。
我要赶紧澄清一下,我个人卑微的科学贡献并不能达到以上所述的地位。然而,正是出于这个原因,我倾向于不将科学与科学“普及”彻底分离。将那些迄今只在专业文献中存在的思想仔细阐述出来,实在是一项困难的艺术。它需要语言上有洞察力的新方法与浅显易懂的比喻。如果你可以强调语言和比喻的新颖,你最终能得到一种新思维。而新思维本身便是对科学的一种原创贡献,正如我之前讨论的那般。爱因斯坦本人便是一位出色的科学普及者。我经常觉得他那些生动的比喻并不只帮助了我们这些读者。它们难道没有为这位极富创造力的天才的思维火花增添燃料吗?
早在20世纪30年代初,从基因角度看达尔文主义的想法已经在费希尔 [1] 和其他新达尔文主义的伟大先驱者的作品中含蓄表达过了。汉密尔顿和威廉斯则在60年代明确表达了这一点。他们的思想使我的思维得到开阔。但我发现他们的表达过于简洁,不够振聋发聩。我坚信一个扩展版本可以使生命万物归位,无论在心灵中还是脑海里。我想要写一本书,赞美基因角度下的进化。它可以集中阐述社会行为的例子,帮助纠正当时盛行的通俗达尔文主义的无意识的类群选择论调的蒙昧。1972年,当时劳资纷争使得实验室停电,我的实验室研究不得不暂停,我便动笔开始写作此书。不幸的是(从某个角度看),大约两个章节完成后,停电结束了。我将这一工程封存,直到1975年我有了一年休假才得以继续。同时这个理论也已经被约翰·梅纳德·史密斯和罗伯特·特里弗斯拓展。我现在可以看到,那是一个神秘时期,所有新思想都在空气中飘浮。我在某种兴奋狂热的状态下完成了《自私的基因》。
当牛津大学找到我出版第二版时,他们坚持认为传统的全面逐页的修订方法并不合适。在他们看来,有一些书显然日后将有一连串的新版本,但《自私的基因》并不是这样的书。第一版有着那个时代的青春气息。当时我们有国外革命的香氛,有一缕华兹华斯的吉祥晨曦。作为那个时代的产儿,若用新发现的事实使其臃肿,或是放任复杂谨慎令其苍老,实在令人扼腕。于是,最初的文本应保持不动,其瑕疵与偏颇也应一并保留。最后的注释则应包括修正、回应与新的发展。全新的章节应当加入,它们的主题在其时代里也将继续带着革命黎明前的情绪。这便是第12与13章。为此我从这些年里最令我激动的两本专业领域内的著作中汲取灵感:罗伯特·阿克塞尔罗德(Robert Axelrod)的《合作的进化》(The Evolution of Cooperation ),因为它给予我们的未来以某些希望,还有我自己的《延伸的表型》,因为它是我这些年的工作成果,也因为——它最有价值的地方是——它可能是我的最佳著作了。
“好人终有好报”的标题出自1985年我参与的BBC电视节目《地平线》。这是一部50分钟的纪录片,由杰里米·泰勒(Jeremy Taylor)制作,以博弈论探讨进化中的合作。这部纪录片的制作,连同另一部来自相同制作人的《盲眼钟表匠》,使我对其职业产生新的敬意。《地平线》的制作人们竭尽全力使自己成为该题目的高级专家(他们的一些节目在美国也能看到,通常以《新星》的名目重新包装)。第12章不仅从中收获了章名,我、泰勒和《地平线》制作组的紧密合作也使第12章的写作获益不少。对此我深表感激。
最近我了解到一个我不敢苟同的事实:一些有影响力的科学家习惯在他们并未参与的作品中署上自己的名字。显然,一些资深科学家要求在作品中署名,只是因为他们贡献了实验场所、科研资金和对文章编辑提出了修改意见。就我所知,他们在学界的声誉可能完全建立于其学生和同事的工作成果之上!我不知道如何与这种不诚实的行为抗争,也许期刊编辑应该要求每一名作者签字表明其贡献。但这不过闲谈而已,我提起这个话题是为了做一个对比。海伦娜·克罗宁(Helena Cronin)对这本书的每一行,甚至每一个字都做了力所能及的改进,却坚持拒绝了成为书中新增部分的共同作者的请求。我对她感激不尽,并对我的感谢必须止于此表示歉意。我还要感谢马克·里德利(Mark Ridley)、玛丽安·道金斯(Marian Dawkins)和艾伦·格拉芬对本书的建议和对一些章节的建设性批评意见。另外还要感谢牛津大学出版社的托马斯·韦伯斯特(Thomas Webster)、希拉里·麦格林(Hilary McGlynn)和其他欣然容忍了我的奇思妙想和拖延的同事。
理查德·道金斯
1989年
[1] 指罗纳德·艾尔默·费希尔(Ronald Aylmer Fisher),英国统计学家、遗传学家。
序言
虽然黑猩猩和人类的进化史大约有99.5%是相同的,但人类的大多数思想家还是把黑猩猩视为畸形异状、与人类毫不相干的怪物,而把人类自己看成通向全能上帝的阶梯。对一个进化论者来说,情况绝非如此。认为某一物种比另一物种高尚是毫无客观依据的。不论是黑猩猩和人类,还是蜥蜴和真菌,它们都是经过长约30亿年的所谓自然选择这一过程进化而来的。每一物种之内,某些个体比另一些个体留下更多的生存后代,因此,这些得以繁殖的幸运者的可遗传特性(基因),在其下一代中的数量就变得更加可观。基因的非随机性的分化生殖就是自然选择。自然选择造就了我们,因此,要想了解我们自身的特性,就必须理解自然选择。
尽管达尔文的自然选择进化学说是研究社会行为的关键所系(特别是同孟德尔的遗传学相结合时),但却一直为许多人所忽视。社会科学领域内一系列研究部门相继兴起,致力于建立一种前达尔文派(pre-Darwinian)和前孟德尔派(pre-Mendelian)的社会和心理世界。甚至在生物学领域中,忽视和滥用达尔文学说的情况一直令人诧异。无论造成这种异常发展的原因何在,有迹象表明,这种状况即将告终。达尔文和孟德尔所进行的伟大工作已为日渐增多的科学工作者所发展,其中著名代表人物主要有费希尔、汉密尔顿、威廉斯和史密斯。现在,本书作者道金斯(Richard Dawkins)把建立于自然选择基础上的社会学说的这一重要部分,第一次用简明通俗的形式介绍给大家。
道金斯对社会学说中这一崭新工作的主要论题逐一做了介绍:利他和利己行为的概念、遗传学上自私的定义、进犯行为的进化、亲族学说(包括亲子关系和社会性昆虫的进化)、性比例学说、相互利他主义、欺骗行为和性差别的自然选择。道金斯精通这一基本理论,他胸有成竹,以令人钦佩的清晰文体展示了这一崭新的工作。由于他在生物学方面造诣颇深,他能够使读者领略生物学文献中丰富多彩和引人入胜之处。凡遇他的观点同已发表著作的论点有分歧时,他的评论就像他在指出我的一处谬误时一样无不一语中的。同时,道金斯不遗余力地把论证的逻辑推理交代清楚,使读者能够运用这种逻辑推理再去扩展这些论据(甚至可以和道金斯本人展开争论)。这些论据可以向许多方面扩展。例如,如果(按道金斯的论证)欺骗行为是动物间交往的基本活动的话,动物就一定存在对欺骗行为的强烈的选择性,而动物也转而必须选择一定程度的自我欺骗,使某些行为和动机变成无意识的,从而不致因极细微的自觉迹象,让正在进行的欺骗行为败露。因此,认为自然选择有利于更准确地反映了世界形象的神经系统这种传统观点,肯定是一种关于智力进化的非常幼稚的观点。
近年来,人们在社会学说方面取得了重大进展,由此引发了一股小小的逆流。例如,有人断言,近年来社会学说方面的这种进展,事实上是为了阻止社会进步的周期性阴谋的一部分,这种进步在遗传上似乎是不可能的。还有,把一些相似而又不堪一驳的观点罗列在一起,使人产生这样一种印象,即达尔文的社会学说,其政治含义是反动的。这种说法同事实情况大相径庭。费希尔和汉密尔顿首次清楚地证明了遗传上性别的均等性。从社会性昆虫得到的理论和大量数据表明,亲代没有主宰其子代的固有趋势(反之亦然)。而且亲代投资和雌性选择的概念,为观察性别差异奠定了客观而公正的基础,这是一个相当大的进展,超越了一般把妇女的力量和权利归结于毫无实际意义的生物学上的特性这一泥潭所做的努力。总之,达尔文主义的社会学说使我们窥见了社会关系中基本的对称性和逻辑性,在我们有了更充分的理解之后,我们的政治见解将会重新获得活力,并对心理学的科学研究提供理论上的支柱。在这一过程中,我们也必将对苦难的众多根源有更深刻的理解。
特里弗斯
1976年7月于哈佛大学
前言
读者不妨把本书当作科学幻想小说来阅读。笔者构思行文着意于引人深思,唤起遐想。然而,本书绝非杜撰之作。它不是幻想,而是科学。“事实比想象更离奇”,暂不论这句话是否有老生常谈之嫌,它却确切地表达了笔者对客观事实的印象。我们都是生存机器——作为运载工具的机器人,其程序是盲目编制的,为的是永久保存所谓基因这种禀性自私的因子。这一事实直至今天仍使我惊叹不已。我对其中的道理虽已领略多年,但它始终使我感到有点难以置信。我的愿望之一是能够凭此使读者惊叹不已。
在写作过程中似乎有3位假想的读者一直在我背后不时地观望,我愿将本书奉献给他们。第一位是我们称之为外行的一般读者。为了他,我几乎一概避免使用术语。在不得已使用专门术语的地方,我都一一详加说明。我不明白为什么我们不把一些学术性刊物里的大部分术语也删掉呢?虽然我假定外行人不具备专业知识,但我却并不认为他们愚昧无知。只要能做到深入浅出,就能使科学通俗易懂。我全力以赴,试图用通俗的语言把复杂艰涩的思想通俗化,但又不丧失其精髓。我这样尝试的效果如何尚不得而知。我的另一个抱负是,让这本书成为一本引人入胜、扣人心弦的读物,使其内容无愧于题材。但这方面我能取得多大成功,心中也毫无把握。我一向认为,生物学犹如神话故事那样迷人,因为事实上,生物学的内容就是神话故事。本书的题材理应激发读者产生莫大的兴趣并带来启发,但我所能做到的充其量不过是沧海一粟,再多我也不敢奢望了。
第二个假想的读者是个行家。他是一个苛刻的评论家,对我所用的一些比拟笔法和修辞手段很不以为然。他总是喜欢用这样的短语:“除此之外……”,“但在另一方面……”,“啧!啧!”我细心地听取了他的意见,纯粹为了满足他的要求,我甚至把书中的一章全部重写了一遍。但归根结底,讲述的方式毕竟还是我的选择。这位专家对我的写作方式恐怕不会完全没有微词吧!但我仍极为热切地希望,即使是他也能在拙作中发现一点新内容,也许是对大家所熟悉的观点的一种新见解,甚至受到启发产生出自己的新观点。如果说我的这种心愿太大,那么,我是否可以希望,这本书至少能为他的旅途消愁解闷?
我心目中的第三位读者是位从外行向内行过渡的学生。如果他至今还没有抱定目标要在哪一方面成为专家,那么我要奉劝他考虑一下我所从事的专业——动物学。动物学固然自有其“实用价值”,且大部分动物又有其逗人喜爱之处,但除此之外,研究动物学有其更为深远的意义:因为宇宙万物之中,我们这些动物当属最为复杂、设计最为完美的“机器”了。既然如此,弃动物学而选择其他学科就令人费解了!对那些已经献身于动物学研究的学生来说,但愿本书能有一定的教育价值,因为他们在学习过程中孜孜不倦钻研的经典理论著作和专业书籍,正是笔者撰写本书的依据。如果他们发现经典理论著作难以理解,那么我的深入浅出的论述,作为入门或辅助材料之类的读物,也许对他们有所助益。
显然,要同时迎合3种类型的读者的口味势必要冒一定的风险。我只能说,对此我始终是十分清楚的。不过,考虑到我的这种尝试所能带来的种种益处,我甘愿冒这种风险。
我是个行为生态学家,所以动物行为是本书的主题。我接受过行为生态学的传统训练,从中获得的教益是不言而喻的。特别值得一提的是,在牛津大学我曾在廷贝亨 [1] 指导下工作过12个年头。在那些岁月里,他对我的影响之深,恐怕连他自己也想不到。“生存机器”这个词语虽非实际出自他口,但说成是他的首创亦不为过。近年来,行为生态学受新思潮的冲击而生机勃发。从传统观点来说,这股思潮的来源不属行为生态学的范畴。本书在很大程度上即取材于这些异军突起的思想。这些新思想的倡导者主要是威廉斯、史密斯、汉密尔顿和特里弗斯,我将分别在有关章节中提及。
各方人士为本书的书名提出过许多建议,我已将他们建议的名称分别移作有关各章的题目:“不朽的双螺旋”,来自克雷布斯(John Krebs);“基因机器”,来自莫里斯;“基因种族”,来自克拉顿——布罗克(Tim Clutton-Brock)和简·道金斯,为此我向他们表示谢意,另外,特向斯蒂芬·波特(Stephen Potter)表示歉意。
尽管假想的读者可以作为寄托虔诚希望的对象,但同现实生活中的读者和批评家相比,毕竟无太大实际意义。笔者有一癖好,文章非改上几遍不肯罢休。为此,玛丽安·道金斯不得不付出艰辛的劳动。对我来说,她对生物学文献中渊博知识的掌握,对理论问题的深刻理解,以及她不断给予我的鼓励和精神上的支持,都是我从事此项工作不可或缺的。克雷布斯也阅读了全书初稿。有关本书的议题,他的造诣比我深,而且他毫不吝惜地提出许多意见和建议。格莱尼丝·汤姆森(Glenys Thomson)和沃尔特·博德默(Walter Bodmer)对我处理遗传学论题的方式提出过既诚恳又严厉的批评,而我所做的修改恐怕还不能完全使他们感到满意,但我希望他们会发现修订后的稿子已有所改进。他们不厌其烦地为我花费了大量时间,对此我尤为感激。约翰·道金斯以其准确无误的眼力指出了一些容易使人误解的术语,并提出了难能可贵的修改意见。我不可能找到比马克斯韦尔·斯坦普(Maxwell Stamp)更适合、更有学问的“外行”了。他敏锐地在初稿中发现了一个反复出现的文体缺陷,这对我完成最后一稿助益匪浅。最后,我还要向牛津大学出版社的罗杰斯表示谢忱。他审阅过我的手稿,所提意见富于助益;此外,在安排本书的出版时,他做了许多分外的工作。
理查德·道金斯
1976年
[1] 指尼古拉斯·廷贝亨(Nikolaas Tinbergen),荷兰动物行为学家与鸟类学家。
第1章 为什么会有人呢?

第2章 复制因子

第3章 不朽的双螺旋
第4章 基因机器

第5章 进犯行为:稳定性和自私的机器

第6章 基因种族
第7章 计划生育
第8章 代际之战

第9章 两性战争
第10章 你为我搔痒,我就骑在你的头上

第11章 觅母:新的复制因子

第12章 好人终有好报
第13章 基因的延伸

第14章 基因决定论与基因选择论
第15章 对于完美化的制约
尾注
下面的注解对应最初的11个章节。每一个注解都对应原文中的一个星号。
第1章 为什么会有人呢?
*1 第2页:……1859年之前试图回答这一问题的一切尝试都是徒劳无益的……
一些人,甚至是那些无神论者,都对辛普森这段引文有着抵触情绪。我承认,当你第一次读到这里时,一定觉得这段话听上去是那么庸俗、粗鲁与狭隘,有点像亨利·福特的名句“历史或多或少都是废话”。但是,抛开宗教性答案不谈(我很熟悉这些答案,所以请节省你们的邮票),当你真正开始回想前达尔文时代对诸如“人是什么?”“生命有意义吗?”“人生目的何在?”等问题的答案时,你所能想到的任何一个答案,除开其本身(一定的)历史价值以外,不都是一文不值吗?有些东西是完全错误的,而所有1859年之前对此类问题的答案都属此列。
*2 第4页:我并不提倡以进化论为基础的道德观……
批评者们时常误以为《自私的基因》是在鼓吹自私乃是我们的生活准则!其他人,可能是因为他们仅仅读了标题或书的前两页,会认为无论我们喜欢与否,自私和其他一些阴暗面是我们天性中不可避免的一部分。如果你像其他很多人那样毫无道理地去认为基因的“决定性”是永远的——绝对而且不可逆,那么你就很容易落入这个错误当中。事实上,基因的决定性仅有统计学的意义(参见41——45页),“晚霞行千里”这句众所周知的谚语是一个很好的类比。从统计上来说,晚霞预示着次日是个晴天,但是我们不会为此下太大的赌注。我们很清楚地知道天气变化多端,会受到很多因素的影响。任何一次天气预报都可能被证明是错误的。那仅仅是一个统计学预报而已。我们并不会认为晚霞就一定代表第二天天气晴朗,那么我们同样也不应该认为基因真能确定些什么。没有任何理由能让我们相信基因的效果不能被其他因素影响甚至逆转。对于遗传决定论的详细讨论,以及为什么会有这些误解,请阅读《延伸的表型》第2章,以及我的论文《社会学:茶杯里的新风暴》(‘Sociobiology: The New Storm in a Teacup’)。我还曾因为声称人类从本质上来说都是芝加哥匪徒而被谴责!但是芝加哥匪徒这个比喻的关键点在于:
了解一个人成功的环境能够告诉你一些关于这个人的事情。这和芝加哥匪徒的哪个具体的品质没有任何关系。我同样也可以用一些其他的类比,例如一个已在英国国教会取得高位的人,或者一个被选入雅典娜神庙的人。在任何一个例子中,我比喻的主体都是基因而不是人。
我在《捍卫自私的基因》(‘In Defence of Selfish Genes’)这篇文章里讨论了这一点以及一些其他字面上的误解。上面那段话就是从其中摘录的。
我必须补充一句,那些本章偶尔出现的政治性对白让我在1989年重新阅读的时候很不舒服。“最近几年里,还需要把这(需要克制自私的贪婪以防止整个组织的灭亡)给不列颠的工人们说多少次呢?”这句话让我听起来就像一名托利党党员。1975年,在我写这句话时,我投票支持的社会主义党政府正尽全力与23%的通胀做斗争,很显然他们会很关注有关高工资的要求。像我这样的评论可以在当时任何一位工党大臣的演讲中找到。现在的英国拥有了一个新右派政府,它将吝啬与自私擡高到了意识形态的高度,我的话因此被联想到带有一些龌龊的意味,对此我表示歉意。但这并不意味着我会收回我说过的话。自私的短见仍然会带来我所提及的那些不想要的后果,但是在现在,如果有人想要在英国找寻关于自私的短见的例子,他不应该将目光瞄准工人阶级。实际上,我也许根本不应该把政治评论这个担子压在科学作品上,因为它们过时得是如此之快。20世纪30年代那些关注政治的科学家的作品在今天都被他们那些过时的讥讽毁掉了,例如那些来自约翰·伯顿·桑德森·霍尔丹及兰斯洛特·霍格本(Lancelot Hogben)的作品。
*3 第7页:……把头吃掉可能反而会改善雄性的性活动。
我是在一个同事所做的关于石蛾的研究讲座中第一次听说这种雄性昆虫的古怪事实的。他提到他希望能在人工饲养环境中使石蛾繁殖,但竭其所能,他始终不能让它们交配。这时候,坐在第一排的昆虫学教授咆哮着说道:“你没有将它们的头去掉?”就像这是一件如此浅显而不应该被忽略的事情一样。
*4 第12页:……选择的基本单位,也就是自我利益的基本单位,既不是物种,也不是群体,严格说来,甚至也不是个体,而是遗传单位基因。
从我写下遗传选择性的宣言开始,我就在重新思考在长期进化的过程中,是否会偶尔出现一种更高层次的选择性。我得赶紧指出,我所谓的“更高层次”和“类群选择性”没有哪怕一丁点儿的关系。我指的是一些更微妙、更有趣的东西。我现在的感觉是,不仅仅某些个体对其他个体有着存活的优势,整个有机体的组合可能也会比其他的组合更容易进化。当然,我们这里提到的进化依然是老版本的进化,通过选择特定基因来完成。因为变异能影响个体的存活率和繁殖率,所以它们仍然受到欢迎。但一个新的在基本胚胎层面的关键变异却可能为今后数百万年的发散变异打开方便之门。一种在胚胎层面的更高层次的选择性使其本身更适应进化:一种朝向可进化性的选择。这种类型的选择性甚至可能积累并发展,而类群选择性达不到这一点。这些观点在我的论文《可进化性的进化》(‘The Evolution of Evolvability’)里有详细论述,它们大部分是当我玩“盲眼钟表匠”时受到的启发,这个计算机程序能够模拟进化的很多方面。
第2章 复制因子
*5 第16页:我的概括性叙述大概与事实不会相去太远。
我们有很多关于生命起源的学说。在《自私的基因》里,我只选择了其中之一来阐述我的主要观点,而不是不厌其烦地全都过一遍。但我并不想留下诸如这是唯一严肃的答案或这是最佳答案这样的印象。事实上,在《盲眼钟表匠》中,我刻意挑选了另一种学说来解释同样的问题,那是凯恩斯——史密斯的黏土学说。在两本书里,我都没有认同所选择的理论假说。如果我再写另一本书,我可能会利用机会尝试从另一个角度来解说,比如德国数学化学家曼弗雷德·艾根(Manfred Eigen)及其同事的理论。我一直以来试图强调的是学说里的那些基本要素。每一个成功的关于任意星球上生命起源的理论都需要有这些基本要素,特别是要有能自我复制的遗传实体这个概念。
*6 第19页:“看哪!一个处女将要受孕……”
一些愤怒的读者致函质问我关于那部神圣的预言书把“年轻妇女”误译为“处女”的事情,并要求我给出答复。伤害宗教感情在现在是一件很危险的行为,所以我最好还是给出一个答复。其实对于那些不能经常在真正学术的脚注中得到满足的科学家来说,这是一件好事。事实上,这一点是研究《圣经》的学者都知道的,也并不会受到他们的反驳。《以赛亚书》中的希伯来词是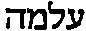 (almah),这个词的含义肯定是年轻妇女,没有任何关于处女的暗示。如果想要表达处女,则应该使用
(almah),这个词的含义肯定是年轻妇女,没有任何关于处女的暗示。如果想要表达处女,则应该使用 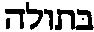 (bethulah,模棱两可的英文单词“maiden”就展示了混淆这两个含义是多么容易)。这个“突变”发生于基督教以前的希腊文《旧约全书》,其中almah被译为
(bethulah,模棱两可的英文单词“maiden”就展示了混淆这两个含义是多么容易)。这个“突变”发生于基督教以前的希腊文《旧约全书》,其中almah被译为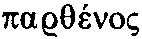 (parthenos),而后者的含义通常是处女。当马太(当然不是那位与耶稣同一时代的使徒,而是福音记录者在其后很久写作而成的)说“这一切的成就,是要应验主借先知所说的话:必有童女怀孕生子,人要称他的名为以马内利”(译自正统英译本)时,他引用了看起来是由希腊文《旧约全书》衍生而来的《以赛亚书》(因为15个希腊单词中只有两个不同)。在基督教学者中,大家广泛接受耶稣为处女所生这个故事是后来篡改的。这大概是由一些说希腊语的信徒带来的,以使这个(误译的)预言看起来是实现过的。在一些现代《圣经》版本中,《以赛亚书》里已经将这个词译为了“年轻妇女”(youngwoman),例如《圣经》英语新译本。在《马太福音》中,“处女”则被正确地保留了,因为在这里它们是由希腊原文翻译而来的。
(parthenos),而后者的含义通常是处女。当马太(当然不是那位与耶稣同一时代的使徒,而是福音记录者在其后很久写作而成的)说“这一切的成就,是要应验主借先知所说的话:必有童女怀孕生子,人要称他的名为以马内利”(译自正统英译本)时,他引用了看起来是由希腊文《旧约全书》衍生而来的《以赛亚书》(因为15个希腊单词中只有两个不同)。在基督教学者中,大家广泛接受耶稣为处女所生这个故事是后来篡改的。这大概是由一些说希腊语的信徒带来的,以使这个(误译的)预言看起来是实现过的。在一些现代《圣经》版本中,《以赛亚书》里已经将这个词译为了“年轻妇女”(youngwoman),例如《圣经》英语新译本。在《马太福音》中,“处女”则被正确地保留了,因为在这里它们是由希腊原文翻译而来的。
*7 第22页:在今天,它们群集相处,安稳地寄居在庞大的步履蹒跚的“机器人”体内……
这段华美的语言(一个罕见的,好吧,比较罕见的溺爱)被幸灾乐祸般地不断引用再引用,用来证明我狂热的“基因决定论”。其部分原因隐藏在对“机器人”一词流行却错误的理解中。我们正处于一个电子学的黄金时代,机器人不再是僵直死板与低能的,恰恰相反,它们已经有了学习能力、智力与创造力。讽刺的是,在1920年卡雷尔·卡佩克(Karel Capek)创造这个单词时,“机器人”其实就是指一个能有人类情感的机器,例如能够坠入爱河。那些认为机器人从定义上就比人类更加“确定”的人都有些思维混乱(除非他们是有宗教信仰的,在那种情况下他们可能会坚持认为比起机器,人类收到了来自神明的关于自由意愿的馈赠)。就像很多对我“步履蹒跚的‘机器人’”这段的批评者一样,如果你也不信教,那么你就该直面接下来这个问题。如果不是一个机器人(尽管这个机器人十分复杂),那么你究竟认为自己是一个什么?我在《延伸的表型》第15——17页讨论了所有这些内容。
这些错误被另一种有效的“变异”积累。正如从神学来看,耶稣由处女所生有其必要性,从恶魔学的角度来看,任何一个称职的“基因决定论者”都必须相信基因“控制”了我们行为的每一个方面也有着相同的必要性。那些基因复制者“创造了我们,创造了我们的肉体和心灵”(22页),这句话像意料中的一样被误引用[例如在由罗斯、卡明(Kamin)和莱文廷所着的《不在我们的基因里》(Not in Our Genes )里,以及之前的一篇莱文廷的学术论文中],“它们控制着我们,身体以及思想”(强调是我加的)。结合我章节的上下文来看,我认为我想表达的显然是“创造”,而这与“控制”风马牛不相及。任何一个人都能看出这点来,这是一个事实,基因并没有像被批评为“决定论”那样强烈地控制着它们的创造物。每一次我们使用避孕措施的时候,我们都轻易地(好吧,相对轻易地)藐视着它们。
第3章 不朽的双螺线
*8 第27页:……某一个基因做出的贡献和另一个基因做出的贡献几乎是分不开的。
这就是我对基因“原子论”的批评者的回答,同样的回答也能在93——96页上找到。严格地说这只是一个预言而非答案,因为其存在早于那些批评!非常抱歉我必须如此完整地引用我自己的文字,但是《自私的基因》中相关的段落似乎太容易被错过了!例如,在《照看团队与自私的基因》(‘Caring Groups and Selfish Genes’,《熊猫的拇指》中的章节)中,史蒂芬·杰·古尔德论述道:
没有基因来“定义”那些毫不含糊的形体特征,例如你的左膝盖骨或手指甲。身体不可能被细分为部件,而每个部件由一个单独的基因负责建造。成百的基因为制造大部分的身体组件做出了贡献……
古尔德在一篇对《自私的基因》的批评中写了这些话。但现在我们来看看我实际的文字(27页):
制造人体是一种相互配合的、错综复杂的冒险事业,为了共同的事业,某一个基因做出的贡献和另一个基因做出的贡献几乎是分不开的。一个基因对人体的不同部分会产生许多不同的影响。人体的某一部分会受到许多基因的影响,而任何一个基因所起的作用都依赖于同许多其他基因的相互作用。
还有(40页):
不论基因在世世代代的旅程中多么独立和自由,但它们在控制胚胎发育方面并不是非常自由和独立的行为者。它们以极其错综复杂的方式相互配合和相互作用,同时又和外部环境相互配合和相互作用。诸如“长腿基因”或者“利他行为基因”这类表达方式是一种简便的形象化说法,但理解它们的含义是重要的。一个基因,不可能单枪匹马地建造一条腿,不论是长腿或是短腿。构造一条腿是多基因的一种联合行动,外部环境的影响也是不可或缺的,因为腿毕竟是由食物铸造出来的!但很可能有这样的一个基因,它在其他条件不变的情况下,往往使腿生长得比在它的等位基因的影响下生长的腿长一些。
在之后一段里我又强调了这一点,拿化肥对于小麦生产的影响类比。这似乎正是古尔德那么确定我是一个天真的原子论者的原因,但是他忽略了我的大段文字,而这些文字正是引出他后来坚持的那种相互作用主义的观点。
古尔德接下来写道:
道金斯将需要另外一个隐喻:基因核心小组,形成同盟,为加入一个盟约的机会而服从,找出大概的环境。
在赛艇的比喻中(42-44页),我已经精确完成了古尔德后来推荐的内容。尽管我们持有如此多的相同观点,但看看这个赛艇的段落,再想想为什么古尔德关于自然选择的主张是错误的吧。他认为自然选择“接受或拒绝整个生物体就是因为所有部件的集合是以一种很复杂的方式相互作用而产生了优势”。而真正关于“合作的”基因的解释应该是这样的:
基因被选择,不是因为它在孤立状态下的“好”,而是由于它在基因库中的其他基因这一背景下工作得好。好的基因必须能够和与之长期共同生活于一系列个体内的其他基因和谐共存,相互补充。(96页)
对基因原子论的批评者,我也在《延伸的表型》书中的第116——117页及239——247页写了一个更加完善的回应。
*9 第32页:我采用的定义来源于威廉斯。
威廉斯在《适应性与自然选择》中的原话是:
我用基因这个词来表达“那个能够以可察觉的频率分离又重组的东西”……一个基因可以被定义为任何一种遗传信息,该遗传信息存在一个数倍于其内部变化速率的有利或不利的选择偏好。
威廉斯的书已经广泛传播,并且被正确地奉为经典,受到许多“社会生物学家”和社会生物学的批评者的重视。我认为很显然,威廉斯从没有认为他自己的“基因选择论”是在支持一些新的或革命性的东西,而我在1976年也同样没有那么做过。我们两个人都认为我们只是又一次地重复那些前人提出的基本原理,例如费希尔、霍尔丹和赖特这些20世纪30年代的新达尔文理论之父所提出的理论。尽管如此,可能是由于我们强硬的措辞,一些人还是很明显地对我们关于“基因是选择的基本单位”这个观点持反对意见,这包括休厄尔·赖特本人。他们最基本的理由是自然选择只能看到整个生物体,而不是它们体内的每个基因。我把对诸如赖特这种观点的回复写进了《延伸的表型》一书中,特别是该书的238——247页。威廉斯最新的关于基因是选择的基本单位这一问题的想法可以在《捍卫进化生物学中的还原论》(‘Defense of Reductionism in Evolutionary Biology’)里找到,新的想法比以往都更加犀利。一些哲学家,例如赫尔(D. L. Hull)、斯特尼(K. Sterelny)和金切尔(P. Kitcher),以及汉普(M. Hampe)和摩尔根(S. R. Morgan)等,最近也对澄清选择的基本单位这一问题做出了有益的贡献。不幸的是,另外的一些哲学家却使其变得更迷惑了。
*10 第38页:……作为遗传单位的个体因为体积太大、寿命太短,而不能成为有意义的自然选择单位……
效仿威廉斯,在我的主张,即一个单独的有机体不能够扮演复制因子在自然选择中的角色中,我在减数分裂上付出了很多琐碎的努力。我现在发现这只是事实的一半,另一半可以在《延伸的表型》的97——99页以及我的论文《复制因子和载体》(‘Replicators and Vehicles’)中找到。如果那些琐碎的努力就是全部的话,一个无性繁殖的个体,例如一只雌性的竹节虫就会是一个真实的复制因子,一种巨型的基因。但是当一只竹节虫发生变化时,比方说少了一条腿,这样的变化是不会传递到后代的。基因则能够世代传递下去,无论是有性还是无性生殖。因此,基因才是真正的复制因子。以无性繁殖的竹节虫为例,整个基因组(它所有基因的组合)是一个复制因子,但竹节虫本身不是。一只竹节虫的躯体并不是前代的复制品。任何一代的躯体都是由卵开始,在其基因组的指挥下全新形成的。而这个基因组才是一个前代基因组的复制品。
所有本书的印刷本都会是一模一样的。它们都是复制品而非复制因子。之所以是复制品,不是因为它们互相复制,而是因为它们都复制于同样一块印版。它们并没有形成一个复制品的世系,没有哪些书是另一些书的祖先。但如果我们复印了一本书的一页,接下来又复印那个复印件,再复印那个复印件的复印件,这样一直下去就创造了一个复制品的世系。在这个书页的世系中,真实地存在着祖先和后代的关系。在这个世系中出现的任何一个新的污点,都会在其后代而非祖先中出现。一个这样的祖先/后代的世系就有着进化的可能。
从表面上来看,连续数代的竹节虫的躯体看上去像是组成了一个复制品的世系,但是如果你实验性地改变该世系中的一员(例如除掉一只腿),该变化不会在世系中传递下去。作为对比,如果你实验性地改变基因组世系中的一员(例如使用X光),该改变则会在世系中传递下去。这些改变,而非那些关于减数分裂的琐碎努力,才是单独个体并不是“选择的基本单位”,也不是一个真正复制因子这一说法的根本原因。大家早已公认拉马克主义遗传理论是错误的这一事实,恰好也是这个事实的重要结论之一。
*11 第44页:另外一种理论为梅达沃爵士首创……
我被指责为什么将这个关于衰老的理论归功于梅达沃,而不是威廉斯(当然,这种指责并不是来自威廉斯本人或他指使的人)。的确,很多生物学家,尤其是在美国,大都是从威廉斯1957年的论文《多效性、自然选择,以及衰老的进化》(‘Pleiotropy Natural Selection and the Evolution of Senescence’)中了解到这个理论的。同样,威廉斯也确实是在梅达沃之前详尽阐述了该理论。尽管如此,我自己的判断是基于梅达沃在1952年的《生物学中一个未解决的问题》(An Unsolved Problem in Biology )及1957年的《个体的独特性》(The Uniqueness of the Individual )中已经提出了该想法的核心概念。需要补充的是,我认为威廉斯对该理论的发展非常有助益,因为他为梅达沃没有明确强调的论断(“多效性”或“多基因影响”的重要性)做出了不可或缺的贡献。汉密尔顿最近在其论文《由自然选择形成的衰老》(‘The Moulding of Senescence by Natural Selection’)中更进一步发展了这类理论。顺便说一句,我收到过很多来自医生的有意思的信件,但我发现没有任何一封来信评论了我关于“愚弄”基因对它们所在躯体的年纪的推测(46——47页)。我仍然不认为这个想法非常肤浅,并且如果这是对的,难道不会很重要吗,比如在医学上?
*12 第47页:性到底有什么益处?
性究竟有什么益处这一问题依然十分的诱人,尽管我们已经有了一些令人深思的答案,比如由盖斯林、威廉斯、梅纳德·史密斯和贝尔(G. Bell)所着的书,以及米肯德(R. Michod)和列文(B. Levin)编辑的一卷书。对于我来说,最激动人心的新想法来自汉密尔顿的寄生虫理论,杰里米·切法斯(Jeremy Cherfas)和约翰·格里宾(John Gribbin)在《多余的雄性》(The Redundant Male )一书中用非技术性的语言解释了该理论。
*13 第49页:……多余的DNA……看作一个寄生虫,或者最多是一个无害但也无用的乘客……
(也见184页)
我认为多余的、未转译的DNA可能是自私的寄生虫。一些分子生物学家接受并发扬了这一观点[参见奥格尔与克里克以及杜利特尔(Doolittle)与萨皮恩扎(Sapienza)的论文],而且用了一个吸引眼球的词语“自私的DNA”。在《母鸡的牙齿和骏马的脚趾》(Hen’s Teeth and Horse’s Toes )一书中,古尔德提出了一个挑衅的(对我!)观点,他认为,不管自私的DNA这一概念的历史源头在哪,“自私的基因和自私的DNA在关于它们的解释的结构上几乎是一样的”。我找到了他在推理上的错误,但有趣的是他也很善良地告诉了我他是怎么找到我的错误的。在一段关于“还原论”与“等级制度”的开场白之后(和往常一样,我并不觉得这个是错的或是有趣的),他继续道:
道金斯的自私的基因在比例上增多是因为它们能够对身体起作用,协助身体应付生存问题。自私的DNA在比例上增多则是出于完全相反的原因——因为它对身体没有任何作用……
我注意到古尔德提到的区别,但我并不认同这就是根本。恰恰相反,我还是认为自私的DNA只是自私的基因整个理论中的一个特例,这也正是自私的DNA这一概念是如何发源的。(比起杜利特尔与萨皮恩扎,奥格尔与克里克引用的第49页那一段,自私的DNA是一个特例这一点,可能在本书的206页更加清晰。顺带说一句,杜利特尔与萨皮恩扎在他们的标题中使用的是“自私的基因”而不是“自私的DNA”)。让我用如下的类比来回应古尔德。那些赋予黄蜂黄黑条带的基因在频率上的增加是因为这种(警告性的)色彩样式强力地刺激了其他动物的大脑。而赋予老虎黄黑色条纹的基因在频率上的升高则是“出于恰恰相反的原因”——因为按理说这种(隐秘的)色彩样式根本不会刺激其他动物的大脑。这实际上是有区别的,与古尔德的区别(在一个不同层面上)很类似。但这是一个关于细节的微妙区别。我们不可能想要去宣称这两个例子“在关于它们的解释的结构上几乎是一样的”。奥格尔和克里克在将自私的DNA与杜鹃的蛋类比时一举击中要害:杜鹃的蛋最终逃脱了检查,因为它们和宿主的蛋看上去几乎是一样的。
另外,最新版的《牛津英语词典》列出了“自私”的一个新的定义,即“对于一个基因或遗传物质,尽管对表型没有任何影响,仍然能够存在或传播”。这是对“自私的DNA”的一个非常简洁的定义,其第二个支持引证也确实关注了自私的DNA。然而在我看来,中间一部分,也就是“尽管对表型没有任何影响”,并不是很好。自私的基因可能对表型没有影响,但很多实际上会有影响。当然词典编纂者也可以说是为了将其含义限制在只用于表示那些不影响表型的“自私的DNA”。但是,他们的第一个支持引证来自《自私的基因》,包含了那些确实影响表型的自私的基因。当然,我也许不应该对被《牛津英语词典》引用这种荣誉吹毛求疵!
我在《延伸的表型》中进一步讨论了自私的DNA(156——164页)。
第4章 基因机器
*14 第55页:就功能而言,我们可以认为大脑和计算机是相类似的……
像这样的陈述很容易困扰那些望文生义的批评者。当然,他们是对的,大脑与计算机有着很多方面的不同。例如,大脑的内在工作方式就和我们用技术搭建起来的计算机有着很大区别。但这并不能削弱我关于它们功能相似的论述。就功能而言,大脑正是扮演了计算机的角色——处理数据、识别样式、短期和长期数据储存、协调操作等等。
当我们提及计算机时,我关于计算机的言论令人可喜地或令人恐惧地变得——看你怎么看——过时了。我在55页中提到“一个脑壳最多也只能塞进几百个晶体管”,但晶体管现在已经被整合进了集成电路。一个头颅里能够装下的晶体管等价物的数量在今天要数以十亿计。我也在58页提到计算机下国际象棋的水平足以媲美一个业余选手。现今,除了一些非常职业的选手,廉价家用计算机上的国际象棋程序已经能胜过所有人了。而世界上最好的程序正在与象棋大师进行一系列的挑战,例如,《旁观者》(Spectator )杂志的国际象棋通讯员雷蒙德·基恩在该刊1988年10月7日出版的杂志上写道:
一位知名选手被计算机击败现在似乎还能造成轰动,但可能不会持续太久了。迄今为止,挑战人类大脑的最危险的金属怪物被古怪地命名为“沉思”,毫无疑问这是在向道格拉斯·亚当斯(Douglas Adams)致敬。“沉思”最近一次出风头是在8月份于波士顿举行的美国公开赛上,它搅得现场人心惶惶。我手头还没有“沉思”的综合评分,该评分应该是对其在瑞士制公开竞赛的成绩的严格测试。但我注意到了它对战伊戈尔·伊万诺夫并取得了非常引人注目的胜利——伊万诺夫曾经击败过卡尔波夫!注意,这也许就是国际象棋的未来。
接下来是我们对该局的逐步重演。下面的文字是基恩对“沉思”第22步的反应:
精彩的一步……它的想法是让皇后占据中心……这个想法直接导致了这一场非常快速的胜利……这个惊人的结局……现在黑方皇后的侧翼已经因该皇后的侵入而被彻底摧毁了。
伊万诺夫对此回复如下:
一次令人绝望的尝试,计算机满不在乎地忽略了……最终蒙羞。“沉思”忽略了皇后的再次提子,转而突然将死……黑棋告负。
除了“沉思”是世界顶尖棋手这一事实,我觉得更令人惊讶的是评论员不得不使用的那些描述人类意识的语言。“沉思满不在乎地忽略了”伊万诺夫的“绝望的尝试”。“沉思”被描述为具有侵略性。基恩评论伊万诺夫的策略时用到了“希望”这个词,但他的语言却昭示着他应该也愿意将“希望”一词用于“沉思”的身上。就个人来说,也许我更希望一个计算机程序能够获得世界冠军。人类需要学会谦虚。
*15 第59页:离我们200光年之遥的仙女座里有一个文明世界。
《仙女座的A》与它的续集《仙女座大爆发》在描述那个外星文明的位置时出现了偏差,究竟是在那异常遥远的仙女座星系呢,还是像我所说的这样,就是在仙女星座其中一个较近的恒星旁?在第一本小说里,那个行星距我们200光年远,当然还在我们银河系内。而在续集中,同样的那个外星文明却被放在了仙女座星系,足足有200万光年远。读者们可以根据你们自己的兴趣来选择相信是200光年还是200万光年,这对我将要描述的故事不会有丝毫影响。
弗雷德·霍伊尔是这两本小说的合著者之一,也是一名卓越的宇航员以及我最爱的科幻小说《黑云》的作者。他在小说中显露的超凡的科学背景与他最近几本和魏克拉玛辛诃合著的书中的废话形成了强烈的对比。他们对达尔文主义的曲解(将其当作一个纯概率的理论)以及他们对达尔文本人恶毒的攻击对证明他们那个吸引人的(尽管不怎么可信)关于星际生命起源的推测毫无帮助。出版商们应该改变这个误解,即一个学者对某一领域的精通意味着他也是另外一个领域的权威。而在这个误解还没有消除之前,知名学者们应该抵御对其滥用的诱惑。
*16 第61页:……战略以及适用于生计的各种诀窍。
对于现在的生物学家来说,用这种战略性的说法来描述一个动物、植物或者一个基因已经很普遍了。试想“把雄性动物视为下大赌注、冒大风险的赌徒,而把雌性动物视为稳扎稳打的投资者”,它们都有意识地提高它们胜出的概率。这类为了方便而选用的比喻通常是无害的,除非落入了那些没有能力理解其中含义的人,或是某些能力过高而只能曲解其含义的人手上,例如,我就找不到任何方法去理解一篇发表在《哲学》杂志上对《自私的基因》的批评文章。这篇文章由玛丽·米奇利撰写,第一句话就基本代表了全文主旨:“基因不可能是自私或不自私的,就像原子不可能嫉妒,大象不可能抽象或饼干不可能有着什么目的。”我在该杂志下一期发表的《捍卫自私的基因》一文是对这篇高度过激的、有恶意的文章的全面回应。看上去一些人似乎多学了一些哲学工具,克制不住想要来展示他们在学术上的存在,尽管这毫无意义。梅达沃对“哲学小说”的评论提醒了我,“哲学小说”能够吸引到“一大群人,常常这些人都是受过良好教育的并且有着学术品位。但这些人却也受了超出他们辩证分析能力的教育”。
*17 第66页:意识的产生也许是由于大脑对世界事物的模拟已达到如此完美无缺的程度,以致把它自己的模型也包括在内。
我在1988年吉福德演讲(Gifford Lecture)《微观的世界》中就讨论了大脑模拟世界这个想法。我依旧不清楚这对我们解决意识本身这个大问题有没有什么帮助,但我承认我很高兴看到它引起了卡尔·波珀爵士的注意,并在达尔文演讲(Darwin Lecture)中提及。哲学家丹尼尔·丹尼特提供了另一个关于意识的理论,推动计算机模拟这个隐喻更进了一步。为了理解他的理论,我们需要先了解两个计算机领域的概念:虚拟机,以及串行和并行处理器的区别。我接下来会先解释清楚这两个概念。
计算机是一台真实的机器,机箱里装着各式硬件。但是在任何一个特定时间,运行程序使得它看上去像是另外一台机器,一台虚拟机器。长期以来对于每一台计算机来说都是这样的,但现代“人机交互”计算机则将虚拟机这个概念生动地带入了每个家庭。在写这段话时,大家公认的交互型计算机市场上的领头羊是苹果的麦金塔。它的成功之处在于,其配备的软件套装使得原本操作困难且不符合人类直觉的机器看起来像是另外一种机器:一台专门为了适应人类大脑和手而设计的虚拟机器。被称为麦金塔用户交互界面的这个虚拟机也可以被看作一台机器。它有着可以按下的按钮,有着像高保真控制台那样可以调节的滑动条。但它是一台虚拟机,那些按钮和滑动条并不是由金属或塑料制成的,它们仅仅是一些屏幕上的图片,你也只是用一根虚拟的手指在屏幕上按下或滑动它们。作为一个人,你感觉能控制它,这是因为你已经习惯用手指来移动物品。我是一个资深的程序员,在25年里用过很多不同的电子计算机,我可以证明使用麦金塔(或其模仿者)与使用之前的计算机有着截然不同的感受。操作该虚拟机,你会感觉到一种无须费力、很自然的感觉,就如同它是你身体延伸出的一部分。该虚拟机使你能够仅仅依靠你的直觉,而无须查使用说明书进行操作。
我现在转向另外一个需要从计算机科学中引入的背景知识,也就是串行和并行处理器的概念。今天的电子计算机大部分都采用串行处理器。它们都有一个计算中心,所有的数据在被处理时都得通过这唯一的电子瓶颈。因为速度非常快,所以它们能够制造出一个能同时处理多个任务的假象。一个串行计算机就像一个国际象棋大师“同时”与二十个棋手对弈,但实际它只是在他们之间不断地轮换。和国际象棋大师不同的是,计算机在不同任务间切换得是如此的迅速与安静,以至于每一个使用者都产生了一个幻想,享受着计算机对自己的单独服务。然而,从本质上来说,计算机只是按顺序对每一个用户进行服务而已。
最近,随着对更高处理速度的要求,工程师们制造了真正的并行处理机器。我最近很荣幸去参观的爱丁堡超级计算机就是其中一员。它包含一个有着数百个“单板机”的并行阵列,每一个单板机都相当于一台现在的台式机。超级计算机的运行方式首先是获取提交的问题,将该问题分解成若干更小且能独立解决的任务,然后再将这些任务分配给单板机组。那些单板机则获得这些小问题,解决然后提交答案并申请一个新的任务。与此同时,其他的单板机组也汇报着它们各自的结果,这样一来,整个超级计算机就能以高于普通串行计算机几个数量级的速度找到最终答案。
我说过一台普通的串行计算机只需要将它的“注意力”在几个任务之间切换得足够快就能够制造出一个就像是并行处理器的假象。我们可以说在串行的硬件之上,存在一台虚拟的并行处理器。丹尼特认为人类大脑所做的恰恰相反。大脑的硬件部分本质是并行的,就像那台爱丁堡超级计算机。而大脑所运行的程序从设计上就是要产生一个串行处理的假象:一台基于并行架构的串行处理虚拟机。丹尼特认为,关于思考的主观影响中最显眼的就是“一件一件的来”,“意识流”,也就是流水般的自我意识。他相信对于多数动物来说,它们并没有这类串行的体验,都是直接使用原本的并行处理的模式。人类的大脑无疑也会直接使用其并行架构去处理很多保持复杂生存机器运转的常规任务。但是,在此基础上,人类大脑同样也进化出一台用来模拟串行处理器假象的程序虚拟机。头脑及其流水般的意识就是一台虚拟机,也就是一个用户友好地与大脑交流的方式,如同麦金塔人机交互界面是一个用户友好地与灰箱子里的计算机实体交互的途径。
当其他物种似乎对它们天然的并行机器满意的时候,为什么人类需要一台串行虚拟机这个问题还没有明显的答案。也许对于人类需要去做的一些更复杂任务有一些更基本的顺序在其中,或者丹尼特错误地将我们特殊化了。他进一步相信串行程序的发展已经成为一大文化现象。同样,这对我来说也不是那么显而易见。但我需要加一句的是,在我动笔之时,丹尼特的论文还尚未发表,我的评论只是基于他在伦敦1988年雅各布森演讲(Jacobsen Lecture)中阐述的信息。我建议读者们在他的论文发表后直接与丹尼特探讨,而非仅仅基于我这不完整的、主观的,可能甚至是润色过的评论。
哲学家尼古拉斯·汉弗莱也曾提出过一套诱人的假说来说明模拟能力的进化是如何导致意识产生的。在他的《内部的眼睛》(The Inner Eye )一书中,汉弗莱举出了一个很有说服力的例子来说明像我们或黑猩猩这样的高等社会性动物都必须变成专业的心理学家。大脑必须处理和模拟世界的很多方面,但是世界的大多数方面与大脑自身相比是如此简单。一个社会性动物生活在一个包含有其他动物的世界,这个世界里有潜在的伴侣、对手、伙伴与敌人。为了能在这样一个世界里存活并取得成功,你必须变得能很好地预测其他个体下一步要做些什么。与在一个社会性的世界里预测下一步要发生些什么相比,要知道一个没有生命的世界下一步要发生什么简直是小菜一碟。那些大学里的心理学家,运用着科学的方法,尚且不能很好地预测人类的行为,社会伙伴们却往往能通过面部肌肉的微小动作及其他一些微妙的线索,令人惊讶地擅长于读心以及猜测行为。汉弗莱相信这种“自然的心理学”技巧在社会性动物中得到了很好的进化,几乎相当于一个额外的眼睛或者另外一个负责分析的器官。这个“内部的眼睛”就是进化过的社会心理学器官,就像外部眼睛是一个视觉器官一样。
到此为止,我觉得汉弗莱的理由还是令人信服的。他接下来主张内部的眼睛是靠自我审视来工作的。每一个动物都审视它自己的感受和情感,借此去理解其他动物的感觉和情感。这个心理学器官依靠自我审视来工作。我不是很确定我是否同意这个观点能够帮助我们理解意识,但汉弗莱确实是一个睿智的作者,他的书很有说服力。
*18 第67页:一个操纵利他行为的基因……
人们有时会为“导致”利他行为或其他一些表面看来很复杂的行为的基因感到不安。他们(错误地)认为在某种意义上行为的复杂度是编码在基因里的。其他基因都是编码蛋白质,怎么会有一个单独的基因来导致利他行为呢?但实际上所谓的基因“导致”了什么只是意味着如果这个基因发生一个变化,那么那东西会跟着产生一个变化。单个遗传性的变化,也就是细胞里遗传分子的某些细节的变化,就会导致原本就很复杂的胚胎发育过程产生变化,因而导致行为的变化。
例如,一个“导致”鸟类兄弟般利他主义的基因突变,一定不是单独地肩负起了一个全新的复杂行为模式的责任。相反,该突变会调节一些已经存在的,而且肯定很复杂的行为模式。这里最有可能的先驱者是家长行为。鸟类通常都会有复杂的神经装置来满足它们喂养和照看它们自己后代的需要。这实际上也是来自它们的祖先,然后经过了很多世代的缓慢的、一步一步的进化。(顺带说一句,对于操纵兄弟关爱的基因心存疑惑者总是前后矛盾的,他们为什么不对同样复杂的关于操纵家长关爱的基因产生怀疑呢?)这个之前就存在的行为模式——本例中的家长关爱——是受一个简单的经验法则调控的,例如“喂养巢里一切张着小嘴喳喳叫的东西”。那么负责“喂养弟弟妹妹”的基因则可以通过加速该经验法则的发育成熟时间来实现。一只携带有兄弟关爱基因突变的幼鸟仅仅是比正常鸟类提前激活其“家长”经验法则。它会把父母巢里的张口喳喳叫的东西——实际是它的弟弟妹妹——当作它自己巢里张口喳喳叫的它自己的孩子。远比不上发明一个崭新的复杂行为,“兄弟行为”可以通过在此前已经存在的行为的发育时间上进行一点小小的变化产生。和通常一样,谬误往往发生于我们对遗传的渐进性的遗忘。事实上适应性进化是通过小的、一步一步的对已经存在的结构或行为的改变产生的。
*19 第67页:卫生品系的蜜蜂
如果这本书的初版就有脚注,那么肯定会有一个脚注来解释那些关于蜜蜂的结论并不是那么完美,就像罗森布勒自己小心翼翼得出的一样。按照该理论,在许多品系中不应该出现卫生品系特有的行为,但是实际上却出现了一个。按罗森布勒自己的话来说:“我们不可能忽略这个结果,无论我们多么想这样做,但我们这个基因假设是基于其他数据的。”一个可能的解释是那个异常的品系发生了一个突变,尽管这并不是那么可信。
*20 第70页:我们可以把这种行为概括地称为联络。
我现在发觉我对这样处理动物联络的方式有些不满意了。约翰·克雷布斯和我在两篇文章里都主张大多数动物的信号都既不是有益的,也不是欺骗性的,更准确地说应该是有操作性。信号是一个动物利用另一个动物的肌肉力量的方式。夜莺的歌声并不包含信息,甚至连一点虚假的信息也没有。它是很有说服力的,催眠并且吸引人的演讲。在《延伸的表型》一书中我写了这类主张符合逻辑的结论,我也在本书第13章里简要介绍了部分内容。克雷布斯和我认为信号是通过我们所谓的读心术和操纵性相互作用而进化来的。关于整个动物信号的另外一个令人震惊的尝试来自阿莫兹·扎哈维。在第9章的一个尾注中,我以比本书第一版相比更加认同的语气讨论了扎哈维的看法。
第5章 进犯行为:稳定性和自私的机器
*21 第79页:进化稳定策略
我现在更喜欢用下面这种简洁的方法解释ESS的关键概念。ESS就是与它的副本能很好相处的一种策略。理由如下:一个成功的策略是在整个种群中占大多数的策略。因此,它就倾向于遇到很多与它一样的副本。除非它能够很好地处理与自己副本的关系,否则它就很难胜出。这个定义没有梅纳德·史密斯的数学定义精确,而且因为这个定义事实上并不完整,所以它也没法取代他的定义。但它的优势在于从直觉上抓住了ESS的基本概念。
比起开始撰写本章之时,以ESS的方式来思考已经在生物学家之间变得更加普遍。梅纳德·史密斯本人在《进化与博弈论》(Evolution and the Theory of Games )一书中总结了截至1982年这一概念的发展。此领域的另一个主要贡献者杰弗里·帕克也写了一则稍微新一点的评论。罗伯特·阿克塞尔罗德的《合作的进化》运用了ESS理论,但是我不会在这里讨论它,因为我的两个新章节之一,《好人终有好报》,主要就是为了解释阿克塞尔罗德的观点而写的。在本书第一版之后,我自己也有关于这个主题的文章发表,名为“好的策略还是进化稳定策略?”(‘Good Strategy or Evolutionarily Stable Strategy?’),还有一篇是接下来会被提及的关于掘土蜂的合作论文。
*22 第85页……还击策略,在进化上是稳定的。
很不幸的是,这个陈述是错误的。在梅纳德·史密斯和普赖斯的原始文献中就有一个错误,而我在本章中重复了一遍。我甚至使这个错误更加恶化,因为我做出了一个很愚蠢的声明,宣称试探性还击策略“几乎”是一个ESS(如果一个策略“几乎”是ESS,那么它实际就不是,进而会被淘汰)。试探性还击策略之所以看上去和一个ESS如此相似,是因为在一群还击策略者中,没有任何其他策略做得更好。但是在一群还击策略者中,鸽子做得一样好,因为其行为和还击策略者的行为无法分辨。因此鸽子才能够插入该种群内。问题就出现在接下来发生的事上。盖尔(J. S. Gale)和伊夫斯(Revd L. J. Eaves)使用计算机模拟了一个动物种群多代的进化。他们发现模拟中真正的ESS其实是一个老鹰与恃强凌弱者的稳定组合。这并不是早期关于ESS的文献经过此类动态处理而发现的唯一错误。另一个很好的例子是我自己的一个错误,我会在第9章的注解中加以讨论。
*23 第86页:遗憾的是,对于在自然界中各种活动所造成的损失以及带来的利益,目前我们知之甚少,不能够给出实际数字。
我们现在已经有了一些很好的关于自然界中利益及损失的野外测量结果,也已经被特定的ESS模型采用。北美的金色掘土蜂的数据就是其中最好的几个之一。掘土蜂并不是我们所熟悉的秋季蜜糖罐上画的群居黄蜂,后者只是为蜂群工作的雌性工蜂。每一只雌性的掘土蜂都是自行其是的。它整个生命都是为了为它的幼虫提供食物以及庇护而存在的。通常来说,一只雌蜂一开始会在树干中打出一个较长的圆孔,其底部则是一个挖空的洞穴。接下来她会出发去捕获猎物(对金色掘土蜂来说猎物就是蚂蚱或长角螽斯)。当它找到猎物时,它会先蜇伤猎物而使其麻醉,再将其拖回自己挖掘的洞穴内。在积累了四到五只蚂蚱之后,它会把卵产在最上面,然后将孔洞封死。卵进而孵化成幼虫,以那些蚂蚱为食。顺带说一句,把猎物麻醉而非杀死是为了防止其腐烂,使其在幼虫食用时仍然是活着的,因此也是新鲜的。正是因为类似的姬蜂也有这种可怕的习性才促使达尔文写道:“我不能说服自己相信一个仁慈且万能的上帝会有意创造姬蜂这样明显故意以活的毛毛虫喂食幼虫的生物……”他也许也可以用法国厨师煮活龙虾以保持其风味作为一个例子。让我们回到那只雌性掘土蜂的故事,它是很独立的,除了在同一区域,还有很多其他的雌蜂也在独自地干着同样的事情,不过其中一些会直接占用其他蜂挖好的洞穴,而不是辛辛苦苦地再挖一个。
简·布罗克曼博士是在黄蜂领域的珍妮·古道尔(Jane Goodall)。她来自美国,和我一同在牛津工作,她带来了汗牛充栋般的数据,记录了两个完整种群生活中的几乎每一个事件,而这两个种群中的每一只雌蜂都有着单独的标记。这份记录是如此的完善,以至于能够很清晰地画出每一只黄蜂个体的时间预算。时间是一个经济学商品:在生活的某一方面花费了较多时间,能花在另一方面的时间就少了。格拉芬把我们叫到一起并教会了我们如何正确地思考时间损耗及繁殖利益。我们在新罕布什尔的种群的雌蜂间发现了一个真实的混合型ESS的证据,尽管我们在另一个密歇根种群里一无所获。大体来说,新罕布什尔的黄蜂们要么挖掘自己的巢穴,要么侵占其他黄蜂所挖的巢穴。根据我们的解释,因为有些巢穴被挖掘者遗弃因此是可用的,所以黄蜂能从侵占中受益。尽管侵占一个巢穴不需要付出太多努力,但侵占者很难知道该巢穴究竟被遗弃与否。那么接下来几天它都会面临双重占有的风险,也许到头来它回来时发现洞已经被封死,它所有的努力就都付之东流了——因为另外那个占有者已经产卵并且获得了所有的好处。如果在一个种群中侵入的数量过多,可用的洞穴就会变得稀少,双重占有的概率也会增高,这样就值得挖洞了。相反,如果许多的黄蜂都在挖洞,大量的洞穴使得侵占变得有利可图。在一个挖掘和侵占差不多有利的种群中会有一个临界侵占比例。如果实际比例低于该临界比例,自然选择倾向于入侵,因为可用的限制洞穴数量十分充足。如果实际比例高于临界比例,可用的洞穴就会产生短缺,自然选择也就会倾向于挖洞。因此,在种群中就维持着一个平衡。细致、定量的证据显示这是一个真实的混合型ESS,每一个黄蜂个体都有挖洞或侵占的可能性,而不是混合了挖洞或侵占专家的一个群体。
*24 第91页:……再精彩不过地展示了这种行为上的不对称性。
有一个来自戴维斯(N. B. Davies)的比廷贝亨关于“留驻者总是胜利”的实验更一目了然的例子。戴维斯研究的是帕眼蝶。廷贝亨的工作成果都是在ESS理论发现之前做出的,而我在本书第一版对ESS的解释都有些马后炮。戴维斯是在ESS理论的启发下开始构思他的蝴蝶研究的。他发现在牛津附近的怀特姆森林中,雄性蝴蝶个体都会保卫太阳的光斑。雌性都受到光斑的吸引,因此光斑就成了一个有价值的资源,值得为之而战。雄性的数量大于光斑的数量,剩余的雄性就在茂密的树冠处等待着它们的机会。通过捕捉雄性,然后再将它们一个一个地释放在一个光斑上,戴维斯发现无论先释放的是哪一只个体,它都会视自己为光斑的主人,而第二个抵达该光斑的则会被视作“侵入者”。毫无例外,所有的侵入者都会迅速地承认失败,让主人拥有对光斑的独自掌控权。在最后一个决定性试验中,戴维斯成功地“愚弄”了两只蝴蝶,让它们都认为自己才是主人而另一只是侵入者。只有在这种情况下,一场重大且长期的战斗才得以爆发。顺便说一下,为了简化问题,在上面所有例子中我都只提到了单独一对蝴蝶,但实际上,这当然是由很多对蝴蝶组成的具有统计学意义的样本。
*25 第93页:似非而是的ESS
詹姆斯·道森(James Dawson)先生在给《泰晤士报》(1977年12月7日)的信中记录了另一个可能看上去是似非而是的ESS例证:“几年时间里,我注意到当一只海鸥将一根旗杆作为制高点时,总是会激起另一只海鸥试图降落其上的欲望,而这和两只鸟的体形大小毫无关系。”
关于似非而是的策略最令人满意的例子来自斯金纳箱里的家猪。该策略和ESS同样稳定,但是我们最好称之为DSS(Developmentally Stable Strategy,发育上的稳定策略),因为这个策略是在动物自己的生存时间内出现的,而非进化意义上的时间。斯金纳箱里的动物会学会自己喂养自己,通过按下一个杠杆,食物就会被自动地运送到食槽之内。试验心理学家很习惯将鸽子或小鼠放入小型的斯金纳箱中,很快小动物们就能够学会通过按下那些精巧的小杠杆来获得食物作为回报。猪也能学会做同样的事,当然是在一个放大版的斯金纳箱中,按压的也是一个非常不精巧的猪鼻杠杆(我在很多年前看过一个相关的研究录像,我清晰地记得当时快要笑死了)。鲍德温(B. A. Baldwin)和米斯(G. B. Meese)利用斯金纳猪圈训练了几头猪,但故事稍微有些不同。他们将猪鼻杠杆放在猪圈的一端,食物分发器放在另外一端。因此,里面的猪就需要先按动杠杆,然后赶紧跑到猪圈的另外一端获取食物,然后跑回杠杆,重复之前的动作。这听上去还不赖,但鲍德温和米斯把猪成对地放入该装置。这样其中一只猪就有机会压榨另外一只了。“奴隶猪”就往返跑着按压杠杆,“主人猪”则坐在食槽旁边享受着刚分发的食物。每一对猪都形成了此类稳定的“主人/奴隶”的关系,一个工作和奔跑,另一个坐享大部分食物。
现在来揭晓为什么似非而是策略中“主人”和“奴隶”的标签全都是颠倒的。每当一对猪达到稳定状态时,最终扮演“主人”或“剥削者”的都是从其他方面看很顺从的那只。而所谓的“奴隶猪”,也就是做了所有工作的那只,是通常强势的那只。任何了解这些猪的人都会做出与之相反的预测,即那只强势猪会当主人,主要负责吃,而另外那只顺从猪应该是少吃多做的奴隶。
这种似非而是是怎么产生的呢?当你按照稳定策略的方式开始思考时,一切都迎刃而解。我们需要做的全部事情只是将时间由进化学尺度降到发育学尺度,也就是两个个体关系发展的时间尺度。“如果强势,就去坐享其成;如果顺从,就去杠杆工作”这种策略听上去好像可行,但不会是稳定的。那只顺从猪也许会按下杠杆,快速地跑回来,却发现那只强势猪的爪子就搭在食槽里,怎么也挪不开。这样那只顺从猪就会很快地放弃按压杠杆,因为得不到一丁点儿回报。但现在想想相反的策略:“如果强势,就去杠杆工作;如果顺从,就去坐享其成。”这种策略将是稳定的,尽管这有着似非而是的结果,那只顺从的猪吃到了大部分的食物。对于强势猪来说,唯一需要达到的要求是当他从猪圈另一端跑回来时食槽内还能剩下一点食物。当它抵达时,它能够毫不费力地把那只顺从猪从食槽旁赶走。只要还有一点食物残渣来回报它,强势猪拉动杠杆的行为就会坚持下去,从而无意中填饱了那只顺从猪。这样那只顺从猪懒惰地斜躺在食槽旁边这一行为便也得到了回报。所以从整体“策略”来看,“如果强势去做奴隶,弱势去做主人”,双方都能够得到回报,该策略就成了一个稳定的策略。
*26 第93页:……某种类型的优势序位。
特德·伯克(Ted Burk)在还是我的研究生的时候找到了进一步的证据证明蟋蟀里存在这类假的优势序位。他还发现一只雄性蟋蟀如果最近刚击败了另一个雄性时,它更有可能对一个雌性示爱。这应该被命名为“马尔博罗公爵效应”,因为马尔博罗公爵夫人的日记中有一篇如此记载着:“大人今天从战场上回来,等不及脱掉马靴就和我缠绵了两次。”《新科学家》杂志的下述报道也许能够提供另外一个名字,该报道描述了雄性激素睾酮水平的变化:“大赛前24小时网球选手的睾酮水平翻倍。赛后,胜利者的睾酮水平维持不变,失败者的则会降低。”
*27 第95页:……ESS概念的发明是自达尔文以来进化理论上最重要的发展之一。
这句话有点过了。我也许有些过激,因为当代生物文献对ESS概念有种普遍的忽视,特别是在美国。例如,这个概念根本就没有在E. O. 威尔逊的巨著《社会生物学》中出现。这种忽视没有持续太长,所以我也能够更加公正地来看待这个问题,而非像个传教士一般。你没有必要非得用ESS语言,只要你想得足够清晰即可。但是ESS语言对你清晰的思考确是一大助力,特别是面对那些详细遗传信息并不明确的案例——而这在实践中占多数。有时候大家会说ESS模型假设繁殖都是无性的。这句话有点容易导致误解,因为无性很容易被假设为有性生殖的反面。ESS模型的真相则是它根本不想让自己去关注遗传系统的细节。与之相反,它们用一种模糊的语调假设相似生相似。对于很多研究来说,这样假设足够了。事实上,这种模糊甚至可能是有益的,因为它能使大脑集中在关键之处,而非那些细枝末节,例如那些在个别案例中根本不可能弄清楚的遗传优势。ESS思考方式作为一个反向思维模板最有用:它能帮助我们避免那些本来可能会使我们误入歧途的理论错误。
*28 第98页:渐进的进化过程与其说是一个稳步向上爬的进程,倒不如说是一系列从一个稳定台阶走上另一个稳定台阶的不连续的步伐。
这个段落很好地总结了现在众所周知的间断平衡理论的一种表达方式。我很内疚地承认,就像当时英国的很多生物学家一样,当我在写我的推测时我完全忽略了该理论,尽管这个理论在3年之前就发表了。那之后,我变得有些急躁,在《盲眼钟表匠》一书中就能看出,我显得有些过分吹嘘间断平衡理论。如果这伤了某些人的心,我表示歉意。如果了解到至少从1976年起,我的心就已经摆正了这一事实,也许能够让这些人感到一丝欣慰。
第6章 基因种族
*29 第104页:……我一直难以理解,为什么一些个体生态学家如此粗心,竟忽略了……
汉密尔顿1964年的文章没有被继续忽视。生态学家们对其从一开始的忽视到后来的承认这段历史使得其成为一个有趣的定量研究材料,一个研究将“觅母”放入觅母库的案例。在我第11章的注释里会追踪这个觅母的发展。
*30 第104页:……我假定我们讲的是整个基因库中一些稀有的基因。
我们假设要讨论的基因在整个种群中是很稀少的,这能方便我们去解释相关性的测量问题,但做这样的假设却有些取巧,然而,实际上汉密尔顿最重要的成绩之一就是证明了无论该基因是稀少还是常见,他的理论都成立。这也是他的理论中大家都很难理解的一个方面。
我们很多人在处理相关性的测量问题时都会产生如下的困惑。一个物种的任意两个成员,无所谓它们是否属于同一个家族,通常90%以上的基因都是相同的。那么,当我们提及兄弟之间的亲缘关系指数是½或第一代堂兄妹之间的亲缘关系指数是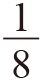 之时,我们的意思是什么呢?正确答案是在任何情况下,在那90%(或其他更准确的数值)的基因以外,兄弟之间还有½的基因是相同的。当然,一个种群的所有个体间也会有一种基础相关性,尽管这种相关程度会比较小。利他主义基因更容易在那些相关性比基础相关性更高的个体中找到,无论这个基础相关性有多大。
之时,我们的意思是什么呢?正确答案是在任何情况下,在那90%(或其他更准确的数值)的基因以外,兄弟之间还有½的基因是相同的。当然,一个种群的所有个体间也会有一种基础相关性,尽管这种相关程度会比较小。利他主义基因更容易在那些相关性比基础相关性更高的个体中找到,无论这个基础相关性有多大。
在第一版中,我通过选取稀有基因这一取巧的做法回避了这个问题。尽管这样做现在看来还没有问题,但始终是不够好的。汉密尔顿自己写到那些基因“在后代中是相同的”,但正如艾伦·格拉芬展示的那样,这句话本身就很令人费解。其他一些作者甚至根本就没有意识到这是一个问题。他们都只是简单地提到相同基因的绝对百分比,而这是一个确定的正向误差。诸如此类的轻率言论导致了一些严重的误解。例如,在对1978年出版的《社会生物学》一书进行激烈抨击的过程中,一位著名的人类学家试图论证如果认真看待亲属选择,那么我们可以预见所有人类都应该互帮互助了,因为所有的人99%的基因都是一样的。我在《关于亲属选择的12个误解》(‘Twelve Misunderstandings of Kin Selection’,这个问题在其中排第5)中简要地回应了这个错误。另外11个误解也值得一看。
在《从几何角度来看相关性》(‘Geometric View of Relatedness’)一文中,艾伦·格拉芬给出了可能是解决相关性测量问题的最终办法。但我不会在这里对其加以阐述。在另外一篇《自然选择、亲属选择和类群选择》里,格拉芬澄清了一个普遍而又重要的问题,即对汉密尔顿“广义适应性”概念的广泛误用。他还告诉了我们关于计算遗传相关性的代价与收益的正确及错误的方法。
*31 第108页:……犰狳……如果有人能到南美去一趟,观察一下它们的生活,我认为是值得的。
关于犰狳我还没有听到过任何前沿进展,但对另外一组“克隆”动物——蚜虫的研究却传来佳音。长期以来,我们都知道蚜虫(也就是青梅子)既可以无性繁殖,也可以有性繁殖。如果你在某个植株上发现了一堆蚜虫,那么很有可能它们都是同一个母本的复制品。而旁边的植株上可能就是另一个母本的复制品了。理论上这种情况很适合亲属选择的利他基因的进化,但直到1977年,人们一直都没有发现蚜虫的利他主义实例。但就在那一年,青木滋之在日本的一个蚜虫种属内发现了一种不育的“士兵”,但那时我已经来不及将其加入书的第一版了。青木在此之后又在数个不同种属里都发现了相同的现象,并且他也有足够证据证明不同的蚜虫种群里至少有四个种群独立进化出了同样的现象。
下面就简要介绍一下青木的发现。蚜虫“士兵”属于一个完全不同的阶级,就像蚂蚁这类传统社会性昆虫里的阶级一样。它们是无法发育成熟的幼虫,因而是不育的。无论是外观还是行为,这些士兵都和它们那些非士兵的同伴不同,尽管它们有着完全相同的基因。士兵通常会比非士兵大:它们有一对超大的前肢,这使得它们看上去就像是蝎子;它们头顶还有一个朝前生长的尖角。它们运用这些武器与可能的捕食者战斗并杀死对方。在整个战斗中,它们很可能会死亡,但就算不死,我们仍然应该正确地认识到它们的遗传利他性,因为它们不可能生育。
回到自私的基因,这里有些什么启示呢?青木并没有清楚地指出是什么决定了哪些个体成为不育的士兵,哪些能够正常繁殖。但我们可以很肯定地说这是由于外部环境而不是基因上的不同。这很显然,因为任何一个植株上的不育士兵和正常蚜虫都携带有完全相同的基因。然而,肯定有一个负责根据环境变化在两种发育路线上切换的基因。为什么自然选择会倾向于这些基因,甚至不管它们中有一些会留在那些不育的士兵体内而不可能向下传递了?因为正是由于那些士兵,同样的基因才能够在可以繁殖的非士兵体内加以保存。这个原因和其他社会性昆虫一样(参见第10章),除了在其他社会性昆虫诸如蚂蚁或白蚁中,不育的“利他者”体内的基因只有统计学上的概率去帮助那些可以繁殖者体内相同的基因。蚜虫的利他者基因之所有享有这种确定性,而不是统计学的可能性,是因为蚜虫士兵和它们为之而战的可以繁殖的姐妹是克隆副本的关系。从某种意义上来说,青木的蚜虫为阐释汉密尔顿理论提供了一个最简洁的活体样本。
那么,蚜虫有资格进入真正的社会性昆虫俱乐部吗?传统上讲,那只是蚂蚁、蜜蜂、黄蜂和白蚁们的阵地,保守的昆虫学家会对其进行百般阻拦,理由例如它们没有一个长寿的老女王。进一步说,作为一个真实的克隆体,蚜虫们并不比你们体内的细胞显得更加的“社会化”。我们可以认为仅有一个的动物在以那株植物为食,只是碰巧它的身体分成了多个蚜虫。就像人体内的白细胞,其中一些蚜虫扮演起了特殊的防御角色。如果继续争辩,“真社会性”昆虫尽管不是同一生物体的一部分,仍然能够展开合作;而青木的蚜虫间的合作仅仅是因为它们都属于同一个“生物体”。我觉得这种文字游戏太无聊了。对我来说,只要你能够了解蚂蚁间、蚜虫间以及人体细胞间发生了什么,你可以随意决定它们是社会性与否。就我自己的喜好,我有充分的理由把青木的蚜虫称作社会性生物体,而非一个生物体的组成部分。因为一个蚜虫个体就能表现出很多单个生物体的关键特征,而一群蚜虫克隆的组合却没有这些特征。这个观点在《延伸的表型》中得到了拓展,相关章节是《重新发现那个生命体》,同样,现在这本书的新章节《基因的延伸》中有相关内容。
*32 第109页:亲属选择肯定不是类群选择的一个特殊表现形式……
类群选择和亲属选择之间的混淆从来就没有消失过,反而可能愈演愈烈。我加倍地想要强调这一点,但有个小小的例外。由于不严谨的措辞,我在本书第一版102页开头留下了一个非常小的谬误。我在初版中写道(这是我这一版少有的几处文字上的改动之一):“我们只是认为第二代的堂兄弟可以接受的利他行为相当于子女或兄弟的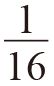 。”正如S.奥尔特曼(S. Altmann)指出的那样,这显然是错误的。这个错误并不是我想要争辩的那个关键。如果一个利他主义动物有一个蛋糕想要分给它的亲戚们,没有任何理由相信它需要给每个亲戚一块蛋糕,也不可能保证蛋糕的大小是由关系的亲疏而决定的。事实上,这听上去可能有些荒谬,但该种群的所有成员,更别提其他种群,都至少是它的远亲。因此,它们每一个都应该来讨要一块仔细称过的蛋糕!与之相反,如果附近就有一个近亲,就没有必要去给远亲哪怕一点儿蛋糕。在其他一些复杂因素诸如回报递减法则的作用下,整个蛋糕都应该给予关系最近的那个亲戚。显然我想要说的是“我们只是预计第二代堂兄弟接收到利他行为的概率相当于子女或兄弟的
”(109页),这也是现在书中所采用的话。
。”正如S.奥尔特曼(S. Altmann)指出的那样,这显然是错误的。这个错误并不是我想要争辩的那个关键。如果一个利他主义动物有一个蛋糕想要分给它的亲戚们,没有任何理由相信它需要给每个亲戚一块蛋糕,也不可能保证蛋糕的大小是由关系的亲疏而决定的。事实上,这听上去可能有些荒谬,但该种群的所有成员,更别提其他种群,都至少是它的远亲。因此,它们每一个都应该来讨要一块仔细称过的蛋糕!与之相反,如果附近就有一个近亲,就没有必要去给远亲哪怕一点儿蛋糕。在其他一些复杂因素诸如回报递减法则的作用下,整个蛋糕都应该给予关系最近的那个亲戚。显然我想要说的是“我们只是预计第二代堂兄弟接收到利他行为的概率相当于子女或兄弟的
”(109页),这也是现在书中所采用的话。
*33 第109页:他有意识地把子女排除在外:他们竟不算亲属!
我表达了我的意愿,希望E. O. 威尔逊能在未来的作品中改变他对亲属选择的定义,也包括把子女算作是“亲属”。我很高兴地在这里汇报——我并不是要说这是我的功劳——但在他的《论人的天性》里,那句有问题的短语,“除了子女以外”,终于被省略了。他补充道:“尽管亲属的定义包含了子女,但当我们使用亲属选择一词时,至少需要其他亲属也受到相关影响,例如哥哥,姐姐,或是父母。”这很不幸地成为现在生物学家通常用法的真实写照,也反映出很多生物学家缺少去理解亲属选择究竟是什么的勇气。他们还是错误地认为亲属关系是深奥且多余的,超越了通常意义上的“个体选择”。但并不是这样的,亲属关系严格遵从新达尔文主义的基本假设,就如同黑夜过后就一定是白天。
*34 第111页:……这样复杂的运算……
亲属选择理论需要动物拥有超现实的计算能力这一说法是错误的。这一谬误在一代又一代的学生中不断地重演,没有一次减弱。犯错的还不仅仅是年轻的学生。如果不是被标榜为对“社会生物学”的“毁灭一击”,由著名社会人类学家马歇尔·塞林斯(Marshall Sahlins)所着的《对生物学的利用和滥用》(The Use and Abuse of Biology )一书还能继续默默无闻地体面存在着。下面引用的是讨论亲属关系是否在人类中同样有效的一段话,这段话好到让人不可能相信它是对的:
强调一点,由于缺少语言学上关于如何计算关系系数r的支持,这个认识论问题成了亲属选择理论的一个重大缺陷。世界上只有极少的语言有分数这个概念,例如印欧语系以及存在于近东和远东的一些古代文明。但对于所谓的原始人类来说,他们根本没有这个概念。猎手和采集者们通常不需要超过一、二、三的计数系统。我都避免去提及更大的问题,动物们该怎么去搞清楚r(自己,第一代堂兄弟)=
这已经不是我第一次引用这个高度揭示性的段落了,我大概也就再引一下自己那无情的回复吧,下面的段落来自《关于亲属选择的12个误解》:
对塞林斯来说真可惜,他居然屈服于诱惑而“避免去提及”动物是如何“搞清楚”r的。他想要嘲弄的这个非常荒谬的观点应该敲响他头脑里的警钟。一只蜗牛的壳是一个优雅的对数螺线,但蜗牛在哪里保管它的对数表呢?因为它眼睛里的晶体缺少对计算折射系数m的“语言学支持”,它又是怎么看对数表呢?绿色植物又是怎么“搞清楚”叶绿素的化学式的呢?
事实上,除了行为学以外,在你能想到的几乎所有生物学的方面,诸如解剖学、生理学,如果按塞林斯的方法你都会走入同样的死胡同。任何动物或植物躯体的发育都需要复杂的数学来精确描述,但这并不意味着动植物自身必须成为一个聪明的数学家。非常高的树木常常会在其树干底部像翅膀一样伸出巨大的板根。对任何一种树而言,树越高,板根就越庞大。大家广泛接受一个事实,即这些板根的形状和大小都接近于能够保持树木直立的最有效方案,然而一个工程师需要非常高深的数学知识才能够证明这一点。尽管树也缺乏相应的数学知识来做出计算,塞林斯或其他什么人也绝对不会来质疑板根背后的理论。那么为什么就要特别地对亲属选择行为提出这个问题呢?这显然不可能是因为这是行为学,而非解剖学。因为塞林斯对很多其他行为学的例子(我是指亲属选择行为以外)都会欣然接受,而不是提出他那个“认识论上的”反对意见。作为一个例子,你可以想象,在某种意义上说,我每接一次球,都需要做出那种经过复杂计算的解释(111页)。人们会情不自禁地想:对于这些很满意自然选择理论的社会学家来说,仅仅由于他们学科历史上一些毫不相干的理由,就会不顾一切地想要专门去找到一些——哪怕一丁点儿——亲属选择理论的问题?
*35 第114页:……我们必须考虑一下,动物在实际生活中是怎样估计谁是它们的近亲的……
我们知道谁是我们的亲属,这是因为别人会告诉我们……
从本书动笔以来,整个亲属选择领域得到了长足的发展。包括我们自己在内,动物似乎都有一些巧妙的辨识亲属的能力,这往往依靠嗅觉。最近一本名为“动物中的亲属识别”(Kin Recognition in Animals )的书总结了到现在为止我们所知道的知识。由帕梅拉·韦尔斯(Pamela Wells)撰写的关于人类的章节指出上面那句话(……我们知道谁是我们的亲属,这是因为别人会告诉我们)需要进行补充:我们至少有强有力的证据证明我们能够运用多种非语言暗号,例如我们亲属的汗味。
对我来说,她开篇的引用就概括了一切:
all good kumrads you can tell
by their altruistic smell
e. e. cummings
除了利他主义,因为一些其他原因亲属们也需要能够认出对方。他们也许也需要在族外婚和族内婚之间找到一个平衡,就如你们将在下一个注释看到的一样。
*36 第114页:……近亲繁殖能产生隐性基因的有害影响有关。(出于某种原因,很多人类学家不喜欢这个解释。)
一个致死基因会杀死它的携带者。就像其他隐性基因一样,一个隐性致死基因也不会产生任何作用,除非当两个碰到了一起。隐性致死基因之所以能够在基因库里存在,是因为大多数个体都只会携带一个拷贝,所以从不会受其影响。任何一个致死基因都是罕见的,因为一旦遇到了另一个致死基因就会导致携带者死亡。但我们的基因组内仍然布满了致死基因,因为它们还可以有很多不同种类。关于人类基因库里究竟潜伏着多少这样的基因还不是很清楚。一些著作认为平均每人体内有两个。如果一个随机的男性和一个随机的女性结合,绝大多数情况下他的致死基因不会和她的致死基因重合,这样一来他们的小孩就没事了。但是如果一对兄妹结合,或者一对父女,情况就大大不妙了。无论我携带的隐性致死基因在整个基因库里是多么罕见,也无论我姐姐的隐性致死基因在整个基因库里是多么罕见,我们俩有着同样隐性致死基因的可能性高得吓人。做做算术,当我和我姐姐结合之后,其结果是对于每一个我携带的隐性致死基因,我们每八个后代中就会有一个生下来就是死的或者早夭。顺便提一下,就基因层面来说,早夭比死胎更加具有致死性:因为死胎并不会过多地浪费父母宝贵的时间和能量。但是,无论你从哪个角度看,近亲结婚都不是一般地有害,它可能是灾难性的。主动乱伦回避的选择压力和任何自然界中产生的选择压力都一样强。
有些人反对达尔文主义对于乱伦回避的解释,但他们可能都没有意识到他们是在反对多么强大的一个达尔文主义案例。他们的辩解有时候是弱得近乎绝望的诡辩。例如,他们通常会说:“如果达尔文主义选择真的让我们从直觉上就反感近亲繁殖的话,我们就不需要去禁止人们这样做了。这样的禁忌之所以存在,就是因为人们还是有近亲繁殖的欲望。因此,反对近亲繁殖的规则并没有‘生物学上的’意义,而它纯粹是‘社会性’的。”这个反对说辞和下面这个很像:“汽车并不需要一个锁去控制引擎开关,因为门上已经有锁了。既然这样引擎锁肯定就不是用来防盗的,它们一定有着纯粹的宗教性的重要性!”人类学家们也很喜欢强调不同的文化有着不同的禁忌,因此就会有不同的关于亲属关系的定义。他们似乎认为这同样也削弱了达尔文主义者想要解释乱伦回避的强烈愿望。但人们或许可以同样认为性欲也不符合达尔文主义的适应性原则,因为不同的文化倾向于不同的性交体位。对我来说,人类中的乱伦回避和其他动物中的一样,都非常可能是很强的达尔文主义选择的结果。
糟糕的事并不仅仅局限于与遗传上和你关系太近的人交配,太远距离的族外婚也同样不好,因为不同支之间会有遗传不亲和性。最佳的中间距离很难准确地预测。你应该和你的第一代堂兄妹结合吗?与第二代或第三代堂兄妹结合呢?帕特里克·贝特森(Patrick Bateson)尝试询问日本鹌鹑倾向于一个怎样的距离。在一个名为阿姆斯特丹装置的实验设备中,研究者让鹌鹑们在迷你橱窗后的一排异性中挑选一个。它们大都挑选了第一代堂兄妹,而非亲兄妹或其他不相干的鹌鹑。进一步的实验显示,年幼的鹌鹑会记住同一窝里同伴的特征,在之后的生活中,会倾向于找具有类似特征的作为性伴侣,但它们不会找过于相似的。
鹌鹑看上去能够避免近亲繁殖,因为它们本身就对一起长大的兄妹们提不起兴趣。其他动物则是依靠遵守社会法则,一个社会性的驱逐法则。例如,青年雄狮会被它的父母从狮群里驱逐,因为它的雌性亲属对它还是很有诱惑力的。仅仅当它占有了另外一个狮群时,它才会交配。在黑猩猩和大猩猩的社会中,则是年轻的雌性去其他群体中寻找配偶。这两个驱逐法则以及鹌鹑的系统都能够在我们自己很多不同的文化中找到踪影。
*37 第119页:由于它们不会受到同一物种其他成员的寄生行为之害……
对于大多数鸟类来说,这也许是对的,但是找到一些会寄生在自己种群的鸟类并不应该使我们太惊讶。其实我们在越来越多的物种里发现了这个现象,特别是现在,新的分子技术能够帮助我们确定谁和谁有关。实际上,自私的基因理论预测这种现象的发生频率要远远高于我们现在所知道的。
*38 第121页:狮群中的亲属选择
帕克和普西(A. Pusey)挑战了伯特伦关于亲属选择是狮群合作的主要推动力这一观点。他们声称在很多狮群中,两头雄狮都是没有关系的。帕克和普西宣称至少互惠的利他主义可以和亲属选择一样作为狮群合作的一个解释。也许两边都是对的。我在第12章主要强调了只有当一开始就有足够临界数量的互惠者,互惠性才能够开始进化。这确保了一个潜在的伙伴有足够的机会可以成为一个互惠者。亲属关系可能是达成这一点最直观的途径。亲戚们自然就容易彼此相像,所以就算是在大的种群尺度上没法满足的临界频率,在家庭范围内仍然有可能得到满足。有可能狮群的合作开始于伯特伦提出的亲属效应,而这个开始就为互惠性的发展提供了必需的条件。这个关于狮群的争议只能够靠事实来解决,而事实从来就只能告诉我们关于一个个例的信息,而不是一个普遍的理论依据。
*39 第121页:如果C是我的同卵孪生兄弟……
现在已经广为人知的是,一个同卵孪生兄弟理论上对你的重要性和你自己一样,只要确定真的是同卵所生。但大家却还不甚了解,对于一个一夫一妻的母亲,她对你也有着一样的重要性,如果你很肯定你的母亲会且仅会继续为你的父亲产下下一代,那么在遗传学价值上来看,你的母亲和你的同卵孪生兄弟,以及你自己,是一样的重要。把你自己想成是一个生殖后代的机器,那么你的母亲就是一个(亲)兄妹生殖机器,而亲兄妹和你自己的后代对你来说有着同样的遗传学价值。当然,这样想缺少了所有现实的考量。例如,你母亲比你年老,尽管这究竟使她比你更适合还是更不适合生育取决于很多情况——我们没法给出一个一般性法则。
这个主张需要假设你能够确保你母亲继续产下你父亲的孩子,而不是其他男人的。这种确信的程度取决于该物种的配偶系统。如果你是一个习惯杂交的物种中的一员,你显然不能确信你母亲的后代就一定是你的亲兄妹。就算是理想化的一夫一妻制,也会出现一个明显的不可逃避的问题致使你的母亲没有你自己合适——父亲可能死亡。无论愿望再美好,如果你的父亲死了,你的母亲也就不可能继续生产他的孩子了,不是吗?
好吧,根据一些事实她可以。这类现象能够出现的事实对亲属选择理论有着很大的意义。作为哺乳动物,我们已经习惯了在交配后经过一段固定且短暂的时间后才生育的概念。一个人类男子可以在死后成为父亲,但不可能是在死后超过9个月的时间(除非有精子库的低温冷藏技术帮忙)。但有许多种昆虫,它们的雌性能够在其体内终生储存精子,一年一年地放出一些来使卵受精。这通常会在它配偶死去很多年以后一直发生。如果你是这些物种中的一员,你可能就能很确定你的母亲会继续成为一个好的“遗传赌注”。一只雌性蚂蚁只会在它的生命早期进行一次婚飞。该雌性在交配之后就会失去它的翅膀,也就永远不会再交配了。尽管得承认,在很多蚂蚁种类里,雌性会在婚飞期间与很多雄性交配,但如果你碰巧是那些雌性始终奉行一夫一妻制的物种中的一员,你完全可以把你的母亲当作至少和你自己一样好的遗传赌注。作为一只幼年蚂蚁与作为一头幼年哺乳动物相比,最好的一点就是你不用在乎你的父亲究竟还是不是活着(事实上,他肯定已经死了)。你可以十分确信你父亲的精子还继续存活着,并且你的母亲还能继续为你生出亲兄妹来。
接下来,如果我们真的对兄妹关怀和诸如昆虫士兵现象的进化起源感兴趣的话,我们需要特别关注那些雌性能终生储存精子的物种。就像第10章讨论的一样,在蚂蚁、蜜蜂和黄蜂的例子中都有一个特别的基因特质——单倍二倍性。这也许预先就使它们更容易变得高度社会化。在这里我想要主张的是单倍二倍性并不是唯一的先决因素。终生储存精子这一习惯可能至少是一样重要的。在理想情况下,这让母亲变得和同卵孪生双胞胎有着同样的遗传学价值,也同样值得“利他主义式”的帮助。
*40 第122页:……社会人类学家或许能够发表一些有趣的议论吧。
这句话现在使我尴尬难堪。我后来才知道,原来社会生物学家对“舅舅效应”不仅有话要说,他们中的很多人多年来不干别的只研究这个!我所“预测”的现象对很多文化都是一个经验事实,而人类学家们几十年前就完全知晓了。进一步来说,当我建议这个特定假说时,即“在不贞行为司空见惯的社会里,舅舅应该比‘父亲’表现出更多的利他行为,因为它有更大的理由信赖同这个孩子的亲缘关系”(122页),我非常抱歉地忽略了一个事实,理查德·亚历山大早就提出过相同的建议。(在本书第一版的印刷后期,我加入了一个脚注说明了这一点)。该假说已经被亚历山大本人以及许多其他人根据人类学文献,使用定量的手法测试过了,也得到了肯定的结果。
第7章 计划生育
*41 第127页:温-爱德华兹……又是这个类群选择论的主要鼓吹者。
比起其他的学术异端,温-爱德华兹的境遇要好多了。尽管犯了一个明显的错误,大家仍然认为他对启发人们认真思考选择原理做出了贡献(尽管我觉得这样做有些过头了)。他自己在1978年豁达地撤回了自己的主张,当时他写道:
现在,理论生物学家达成共识——想要做出一个可信模型让慢吞吞的类群选择打败迅猛的自私基因是不现实的,那些迅猛的自私基因能够带给每一个个体好处。因此我接受他们的意见。
尽管第二个想法看上去如此豁达,但很不幸他有了第三个想法:在他新书里又反悔了。
由于那些众所周知的原因,类群选择理论在现在比我第一版书出版时更不受到生物学家的青睐。请你不要介意地想想相反的场景:新生代已经出现,特别是在北美,他们到处散播“类群选择”一词,就如同在为新娘撒上五彩花瓣。这个词被随意地用到一些其他事物上,而这些事物原本(对我们剩下的人来说现在也是)就是另外一些东西,例如亲属选择。我知道对这种语言上的暴发户感到困扰是没有意义的。但是,关于类群选择的所有问题早在10年前就被约翰·梅纳德·史密斯完美解决了,所以现在发现我们两代人之间,两个国家之间,就因为这一常用语言而分隔开来让人很气恼。更加不幸的是,那些姗姗来迟的哲学家使得这个本来就很混乱的术语变得更加混乱。我推荐艾伦·格拉芬的《自然选择、亲属选择和类群选择》,这是一个将新类群选择相关问题梳理仔细、思路清晰的著作,我也希望这最终能解决混乱的问题。
第8章 代际之战
*42 第143页:1972年,特里弗斯……巧妙地解决了这个难题……
特里弗斯在20世纪70年代发表的一系列论文是我在写本书第一版时得到的最重要的启发之一。他的观点也在第8章里占据了重要地位。他总算也出版了自己的书,《社会的进化》。我推荐这本书,并不只是因为其内容,这本书的风格也很好,在保证思路清晰、学术正确的同时,又有着一点玩世不恭的态度,还掺杂些许自传式的独白。我情不自禁地先要引用这样一句,该句是那么独特。特里弗斯在肯尼亚观察到两只处于敌对关系的雄性狒狒时,他用这样的文字来表达他的激动之情:“还有一个原因使我这么激动,这是一次无意中对亚瑟做出的观察。亚瑟是一个男子汉气概十足的一流年轻雄性……”特里弗斯关于亲代——子代冲突的新章节使整个领域焕然一新。实际上,除了一些实际例子,他1974年的文章还真不用加什么了。该理论经受住了时间的考验。更加详细的数学和遗传学模型已经证实了,特里弗斯那大部分只是语言的论证完全符合现在人们接受的达尔文主义理论。
*43 第156页:按他的说法,亲代总归占上风。
在他1980年的著作《达尔文主义及人类事务》(Darwinism and Human Affairs ,第39页)中,亚历山大勉强承认他的主张是错误的,他声称在亲代——子代冲突中亲代获胜是完全符合基本达尔文主义假设的。他的毕业论文描述亲代在世代争斗中相对于它们的子代有着不对称的优势。但是现在在我看来,这一点也许可以通过另外一个不同的论点来支撑,该论点是我从埃里克·恰尔诺夫那里得到的。
恰尔诺夫当时正在写一些关于社会性昆虫以及不育阶层的起源方面的东西,但他的论据的作用其实更广泛,所以我要将其运用到一般的情况上来。试想一个一夫一妻制种群里的一只即将成年的幼年雌性,并不一定要是昆虫。它现在会面临一个两难境地,究竟应该离开然后试着生育自己的后代呢,还是留在父母的巢穴里帮忙照看更年幼的弟弟妹妹?基于该物种的繁殖习惯,它能很确定自己母亲会长期继续生育它的亲弟弟和亲妹妹。根据汉密尔顿的逻辑,这些弟弟妹妹对它的遗传学价值和它自己的后代是一样的。既然考虑到了遗传学关系,那这只年轻的雌性就应该不再纠结于这两种选择了;它不关心究竟是走还是留。但是它的父母,则会很关心究竟它做出怎样的决定。从它年长的母亲的角度来看,这个选择决定了它是有更多儿女还是孙辈。新的儿女的价值从遗传学上来说比孙辈要高一倍。如果我们把子代究竟是走还是留下帮助父母看作一个亲代与子代的冲突的话,在这场冲突里亲代显然很容易最终获胜,因为只有它们视其为一个冲突。
这就像两个运动员之间的一场较量,其中一个运动员如果赢了就能够获得1 000英镑的奖励,而他的对手无论输赢都能得到1 000英镑。我们可以预测到前一个选手会更加努力,如果两人本来就实力相当的话,那他很可能会赢。其实恰尔诺夫的观点比这个比喻还要强得多,因为无论他们的收益如何,全力以赴地进行一次比赛对很多人来说并不是一个很高的代价。那样一个理想化的奥林匹克对于达尔文游戏来说实在太奢侈了,在达尔文游戏里,在一个方面做出的努力就意味着在另一个方面会有所欠缺。就像是你在一场比赛中投入过多精力的话,你就很难赢得下一场比赛了,因为那时你已精疲力竭。
随着物种变化,条件也会有所不同,因此我们很难每次都预测出达尔文游戏的结果。但是如果我们只考虑最亲的基因相关性,并且假设一个一夫一妻的配偶制度(这样女儿才能够确信它的弟弟妹妹们是亲弟弟妹妹),我们可以推测出年长的母亲会成功地让它刚成年的女儿留下来帮忙。母亲会得到一切,而女儿自己不会有任何动力去反对母亲,因为对它而言,手上的选择并没有什么区别。
再一次强调,这是一个“其他条件一样”类型的主张。尽管其他条件经常都不会完全相同,恰尔诺夫的推论对于亚历山大或其他任何一个支持亲代操纵学说的人还是会有帮助。在任何情况下,亚历山大的实际论断都是会被采纳的——亲代要大一些、强壮一些等等。
第9章 两性战争
*44 第162页:……那么彼此毫无血缘关系的配偶的利害冲突会激烈到何种程度呢?
像往常一样,开篇这句话隐含了“其他条件一样”这一前提。配偶显然很可能从合作中获得巨大收益,这会在本章一遍又一遍地出现。而且,配偶很可能是在进行一场非零和博弈,双方都能在合作中受益,而并非其中一个的获益就是另一个的损失(我会在第12章解释这个观点)。这也是本书中一个我过分着墨于生活中怀疑、自私一面的地方。在当时这看上去是有必要的,因为那时关于动物求偶的主流看法远远地倒向了另外一面。人们近乎普遍地抱持着一种毋庸置疑的假设,即配偶会毫不吝啬地互相合作,从未考虑过互相利用的可能性。在这种历史背景下,我这种看起来很阴暗的开篇语就可以理解了,但现在我会采取更加温和的语调。相似地,在本章最后我做出的关于人类性别角色的评论现在在我看来其语言也有些幼稚了。有两本书更深入地探讨了人类性别的进化,分别是马丁·戴利(Martin Daly)和马戈·威尔逊(Margo Wilson)的《性、进化与行为》(Sex , Evolution , and Behavior )以及唐纳德·西蒙斯(Donald Symons)的《人类性趣的进化》(The Evolution of Human Sexuality )。
*45 第164页:……但雄性个体可以繁殖幼儿的数量实质上是无限的,这就为雌性个体带来了利用这种条件的机会。
现在来看,强调精子和卵子的大小区别就是性别的基础似乎有些误导作用。就算一个精子小而廉价,但要制造百万计的精子并成功地在所有的竞争条件下将它们注入雌性体内并不是一件轻松的事。我现在倾向于用如下方法来解释雄性和雌性之间的基本不对称性。
假设我们有两个性别,它们并不包含雄性或雌性的特征。我们用一种中性的方式将它们命名为A和B。唯一需要注明的是任何一次配对都必须发生在A和B之间才行。现在,任何一只,无论A还是B,都面临这一个抉择:用于与对手战斗的时间和精力就没办法用来照顾后代了,反过来也一样。它们俩中任何一个都需要在这两个矛盾的选择中找到一个平衡。我接下来要讲到的观点就是,A们可能会与B们找到两个不同的平衡点,一旦达到,它们就很可能有了一个逐渐变大的区别。
假设两种性别的动物,也就是A们和B们,从一开始就有了不同。一个通过在后代上投资而获得成功,另一个则是在战斗(我用“战斗”一词来指代所有同性间的竞争)中付出努力而得到成功。一开始两性之间的区别还十分微小,因此我的观点就是存在一种固有的趋势使这种区别变大。好比A一开始由战斗带来的繁殖优势就比亲子行为带来的优势大,而B恰恰相反,亲子行为在它们这儿能带来比战斗稍微高一点的繁殖优势。举个例子来说,这就意味着尽管A当然也能从照顾后代中获益,但A中一个成功的照看者和一个不成功的照看者之间的区别,会小于A中一个成功的战士和不成功的战士之间的区别。在B群体中,则是完全相反的。因此,对于一定量的付出来说,A会通过努力战斗来提高自己的优势,而B更可能把注意力从战斗转向照看后代。
因此在接下来的世代中,A们就会比它们的父母们战斗得更多一些,而B会比它们父母们战斗得少些而多花一些时间照看后代。战斗能力最强的和最弱的A之间的区别将变得更大,而照看能力最强的和最弱的A之间的区别将变得更小。这样一来,通过在战斗方面付出努力,A可以得到更多收益,而照看后代的收益变得更小了。随着世代更替,相反的事情也发生在B的群体中。这里的关键点是两性间一个微小的初始差别能够自我强化:选择可以发源于一个初始的、微小的不同,然后将其慢慢地变大,再变大,直到A变成了现在我们所说的雄性,B则是我们所说的雌性。那个初始区别小到可以随机产生。当然,两性的初始条件也不大可能是一模一样的。
你可能会注意到,这个论点和那个关于原始配子早期分化出精子和卵子的理论很相似。该理论由帕克、贝克和史密斯首先提出,164页也有相关讨论。刚才所讲的论点更加普通一些。分化出精子和卵子只是基本性别分化的一个方面。与把精子与卵子分化作为根本,然后将所有雄性与雌性的特征都归因其上相比,我们现在有了一个能以相同方式去解释精子与卵子分化以及其他方面分化的理论了。我们只需要假设有两个需要互相配对的性别,我们不需要知道关于那两个性别更多的信息。由这个最小化的假设开始,无论一开始两性是多么相同,我们都积极地期待它们会分化成两个专攻于相反且互补的生殖技能的性别。精子和卵子的分化只是这个更加一般的分化的一个征兆,而不是其原因。
*46 第173页:让我们采用史密斯用以分析进犯性对抗赛的方法,把它运用于性的问题上。
我们的意图是要在一个性别内找到一个进化稳定的混合策略,并与另外一个性别的进化稳定的混合策略相匹配。梅纳德·史密斯自己又进一步地发展了这个观点,而艾伦·格拉芬和理查德·西布利(Richard Sibly)也独立地做了差不多的事。格拉芬和西布利的论文在理论上要更高深一点,而梅纳德·史密斯的更方便用文字解释。简要地说,他开始考虑两个策略,坚守(Guard)和离弃(Desert),供两性任意选择。正如我“羞怯/放荡和忠诚/薄情”的模型一样,有趣的问题在于,雄性的哪种策略组合与雌性的哪种策略组合在一起是稳定的?答案取决于我们对该物种特定经济状况的假设。有趣的是,无论如何改变经济状况假设,我们都得不到一个整体连续的定量变化的稳定解。该模型倾向于落入四个稳定解之一。这四个解都依据符合条件的物种来命名。它们是鸭型(雄性离弃,雌性坚守)、棘鱼型(雌性离弃,雄性坚守)、果蝇型(双方离弃)以及长臂猿型(双方坚守)。
还有些更有趣的东西。还记得第5章里ESS模型可以在两种结果之间任选其一,且两者同样稳定吗?好吧,这对于梅纳德·史密斯模型来说也是正确的。其中最有趣的一点是,对比其他策略对,那个满足这种结果的策略对在同一个经济情况下保持着共同稳定。举例来说,在一组情况下,鸭型和棘鱼型都是稳定的。究竟两个中哪一个最终胜出取决于运气,或者说得更准确一点,取决于进化历史上的偶发事件——初始条件。在另外一组情况下,果蝇型和长臂猿型则是稳定的。同样,在任何一个物种中,都是由历史偶然性来决定究竟哪一型最终胜出。但是没有任何情况能使长臂猿型和鸭型共同稳定,也没有哪种情况能使鸭型和果蝇型共同稳定。对亲和与不亲和的ESS进行这种“稳定对”(表示两种策略的组合)的分析能为我们重建进化历史提供一些有意思的结论。例如,它使我们相信在进化历史中,配偶系统中某些特定转换是可能的,其他的则不可能。梅纳德·史密斯通过简要枚举动物界的配偶方式来探索这些历史上的关系,其结论是这样一个令人难忘的反问:“为什么不是雄性哺乳动物负责哺乳呢?”
*47 第176页:但实际上,像那种情况一样,不存在任何摇摆现象,这是能够加以证明的。整个体系能够归到一种稳定状态上。
我很抱歉这个论点是错误的。然而,这个错误很有意思,所以我在原文保留了这个错误,现在花一点时间来讲解一下。这和盖尔和伊夫斯指出的在梅纳德·史密斯和普赖斯的原始文献中的问题如出一辙。我这个错误则是由在奥地利工作的两位数学生物学家指出来的。他们的名字是P. 舒斯特(P. Schuster)和K. 西格蒙德(K.Sigmund)。
我已经正确地计算出了雄性中忠诚和薄情的比例,以及雌性中放荡和羞怯的比例。这些比例能满足两种类型的雄性都同样成功,两类雌性亦然。这实际上就是一个平衡态,但是我没有去检查这是否是一个稳定平衡。这个平衡态有可能更像是一个危险的刀锋,而不是一个安稳的峡谷。要检查其稳定性,我们就要看当平衡态被人为轻微扰动后会发生什么(推动一个立于刀锋上的球,你就再也找不到它了;推动一下峡谷中的球,它还会自己回来)。在我那个有着特定数值的例子中,雄性的平衡比例为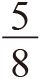 忠诚和
忠诚和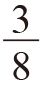 薄情。现在,如果群体中薄情者的比例随机升高到一个比平衡态稍高的数值会发生什么呢?为了使这个平衡态满足稳定及自我修正的标准,薄情者必须马上开始有稍差一些的表现。很不幸,正如舒斯特和西格蒙德指出的那样,这并不是实际情况。恰恰相反,薄情者开始变得更好了!它们在群体中的比例远非自我稳定,而是自我加强的。它们的比例会继续增高,但不是无限的,而是到某个点为止。如果你在计算机上动态模拟该模型,就像我现在所做的那样,你会得到一个无限重复的循环。具有讽刺意味的是,这和我在176页上假想的那个循环一模一样,但当时我觉得我只是用它来作为一个解说工具,就像鹰和鸽子一样。通过与鹰和鸽子类比,我非常错误地认为那个循环只是一个假说,也错误地认为该系统会真的达到一个稳定平衡。
薄情。现在,如果群体中薄情者的比例随机升高到一个比平衡态稍高的数值会发生什么呢?为了使这个平衡态满足稳定及自我修正的标准,薄情者必须马上开始有稍差一些的表现。很不幸,正如舒斯特和西格蒙德指出的那样,这并不是实际情况。恰恰相反,薄情者开始变得更好了!它们在群体中的比例远非自我稳定,而是自我加强的。它们的比例会继续增高,但不是无限的,而是到某个点为止。如果你在计算机上动态模拟该模型,就像我现在所做的那样,你会得到一个无限重复的循环。具有讽刺意味的是,这和我在176页上假想的那个循环一模一样,但当时我觉得我只是用它来作为一个解说工具,就像鹰和鸽子一样。通过与鹰和鸽子类比,我非常错误地认为那个循环只是一个假说,也错误地认为该系统会真的达到一个稳定平衡。
舒斯特和西格蒙最后的不满言论说明了一切:
简要来说,我们能得出两个结论:
(a)两性之间的争斗与捕食之间有着很多共同点;以及
(b)情侣之间的行为就像月亮一样有着盈亏变化,并且像天气一样难以预测。
当然,人们在此之前并不需要用微分方程才能计算出这点。
*48 第179页:……这种父方的献身精神……在鱼类中却很常见。这是为什么呢?
塔姆辛·卡莱尔本科时所做的这个关于鱼的假设现在由马克·里德利进行了比较测试,他彻底审视了整个动物界的亲代照看行为。就像卡莱尔的假说一样,他的论文也是一个伟大的创举,而且同样也是起始于本科时写给我的一篇短文。不幸的是,他没能找出有利论据支持该假说。
*49 第182页:……某种不稳定的、失去控制的过程……
费希尔只是非常简要地描述过他关于性选择的失控理论,现在该理论已经由R.兰德(R. Lande)和其他人数学化了。这已经成为另外一个学科,但如果有足够的空间,该理论还是可以用非数学术语来解释的。但这需要整整一章来解释,所以我的《盲眼钟表匠》(第8章)就专门叙述了这个问题,因此我在这儿就不用赘述了。
相反,我会在这里阐述一个我从未在任何书中充分强调的关于性选择的问题。自然是怎样维持进化所需要的变种的?达尔文选择只有当存在足够基因变种的情况下才可能发生。比方说,你能够成功地繁育一种拥有更长耳朵的兔子。野外种群中的大多数兔子都会有一对中等大小的耳朵(这是根据兔子的标准描述的;当然以我们的标准兔子的耳朵都是非常长的)。一部分兔子会有着比平均值稍短的耳朵,一部分则更长一些。通过选取那些拥有最长耳朵的兔子进行杂交,你就能成功地增加它们后代的平均耳朵长度。但如果你继续在这些长耳兔中选择最长耳朵的进行杂交,终有一天你会发现需要的变种已经不复存在了。它们都拥有了“最长”的耳朵,进化会渐渐陷入停滞。在一般的进化中这种情况并不是一个问题,因为大多数环境都不会在同一方向上施加如此稳定而且毫不动摇的压力。对动物身体任何一部分来说,最好的长度通常都不意味着“无论平均值是什么,都要比平均值更长一些”。最好的长度一般都是一个固定值,比如3英寸。但是性选择却真的有这种追逐永远达不到的最优值的难题。雌性的偏好可能真的是去追求更长的雄性耳朵,无论现在该种群的耳朵已经多长了,这样变种可就真的不够用了。但性选择看上去似乎真的有效,因为我们确实能够发现那些荒诞夸张的雄性装饰物。我们似乎面临一个矛盾,我们可以将这个矛盾称为变种消失悖论。
兰德认为要解决这个矛盾得靠突变。他认为始终都会有足够的突变可以让选择继续进行。人们之前之所以对这种说法存疑,是因为对于一次一个基因来说,任意一个基因位点突变的概率都小到不足以解决变种消失悖论。兰德提醒我们,“尾巴”或是任何一个被性选择的东西都是受到不限定数量的众多不同基因的影响,最终结果都是由这些小的影响叠加而成的。更进一步说,随着进化的前进,这些相关联的不同基因的组合也会发生变化:新的基因会被加入这个影响尾巴“长度变种”的集合,旧的则会被淘汰。变异可以影响这个大型的动态变化基因集中的任何一个基因,于是变种消失悖论就自己消失了。
汉密尔顿对这个矛盾的解释有些不同。他用他如今回答大部分问题的方式来回答:“寄生虫理论”。回想一下兔子耳朵。兔子耳朵的最佳长度大概取决于很多声学因素,没有任何理由相信这些因素会随着世代更替毫不停息地向着同一个方向变化。兔子耳朵的最佳长度可能不是一个绝对的数值,但是选择仍然不可能让其向某一特定方向发展,甚至迷失于现有基因库能够轻易产生的变种范围之外。因此,根本没有变种消失悖论。
但现在来看看由寄生虫带来的剧烈波动的环境。在一个充满寄生虫的世界里,会有一个倾向于对其产生抵抗力的强大选择压力。自然选择会青睐那些最不易受到碰巧就在身旁的寄生虫伤害的兔子个体。关键点是,可能并不止那一种寄生虫出现。瘟疫来了又去,现在可能是黏液瘤病,下一年就有可能是兔子中的黑死病,再下一年是兔子的艾滋等等。然后,也许是一个10年的循环,又轮到了黏液瘤病,接着继续。或者是黏液瘤病毒自身进化出了能够破解由兔子经过逆向适应而得到的抵抗能力。汉密尔顿试想了这种逆向适应和逆向——逆向适应的无休止的循环,该循环倔强地对“最佳”兔子的定义进行不断更新。
所有这些要点在于,在疾病抵抗的适应和对现实环境的适应之间有一些很重要的区别。尽管对兔子的腿长可能有一个确定的“最佳值”,但是若从疾病抵抗的角度来说,是不存在一只明确的“最好的”兔子的。随着当下最危险疾病的改变,当下“最好的”兔子也会随之发生变化。寄生虫是唯一这样做的选择压力吗?比如说,捕食者与猎物又怎么样呢?汉密尔顿承认那和寄生虫很像,但它们并不像很多寄生虫一样进化得这么快,而且寄生虫比起捕食者与猎物更容易进化出具体到基因对基因的逆向适应。
汉密尔顿把寄生虫带来的循环挑战改造成为了一个大一统理论的基础,即他关于究竟为什么性选择会存在的理论。但这里我们只关心他用寄生虫来解决性选择中的变种消失悖论。他相信雄性间可以遗传的疾病抵抗才是雌性选择它们的最重要因素,因为疾病是如此恐怖,如果雌性能够有办法在潜在的配偶中找出患病个体,那么它们就会有巨大的优势。一个表现得像一个很好的诊断医生的雌性,只会选择最健康的雄性作为配偶,因而更可能让它的孩子获得健康的基因。现在,因为对“最佳兔子”的定义在不断变化,当雌性观察雄性时,就必须得有些重要特征让它们做出选择。总是会存在一些“好的”雄性和一些“差的”雄性。他们在几次选择之后不可能还都是“好的”,因为到时候寄生虫已经发生了变化,因而关于“好”兔子的定义也发生了变化。能抵御某一株黏液瘤病毒的基因并不能很好地抵御由变异带来的下一株黏液瘤病毒。瘟疫会这样不断地循环、进化,毫不停歇。寄生虫从不停止,所以雌性们也不能停下它们不懈寻找健康配偶的脚步。
当雄性被像医生一样的雌性仔细检查时,它们会如何反应呢?那些假装健康的基因会取得优势吗?可能一开始是的,但选择会导致雌性加强它们的诊断技巧,从健康个体中排除那些冒牌货。到最后,汉密尔顿相信雌性的诊断技巧会变得足够敏锐,以至于雄性不得不开始为它们的诚实做广告(如果它们真的做广告的话)。如果任何一个性广告在雄性中变得过于夸张的话,这一定是由于其本身就是一个健康的真实指标。雄性会进化得让雌性更容易看出它们是健康的——如果它们真的是的话。真正健康的雄性会很高兴宣扬该事实。不健康的个体当然不愿意,但它们又能做什么呢?如果它们不尝试去展示健康证书,雌性们肯定会对其得出最坏的诊断结论。顺带说一句,这里提到医生并不意味着雌性会对治愈雄性感兴趣,它们只对诊断感兴趣,并且这也不是一个利他主义的兴趣。我也假设我不需要为诸如“诚实”和“得出结论”这样的比喻道歉。
回到广告这一点上,这就有点像雄性被雌性逼着进化出了一个永远插在它们嘴里、能够让雌性清晰读数的医用温度计。这些“温度计”会是怎样的呢?好吧,想想雄性极乐鸟那不可思议的长尾巴。汉密尔顿的解释总体上来说更实在一些。鸟类的一个常见症状是腹泻。如果你有一根长尾巴,腹泻就很有可能弄脏它。如果你想要掩盖你被腹泻困扰这一事实的话,最好的办法就是避免拥有长尾巴。同理,当你想要广而告之你不存在腹泻这一事实的时候,最好的办法就是长出一个非常长的尾巴。那样的话,你尾巴很干净这一事实就变得更加引人注目。如果你根本就没什么尾巴,雌性们看不出它究竟干净与否,就只能得出最坏的结论。汉密尔顿可能不愿意自己来做出这个特定的关于鸟类寄生虫尾巴的解释,但这是他喜欢的解释方式之一。
我将雌性比作诊断医生,而雄性通过到处安放“温度计”来简化前者的工作。想一下医生用的其他一些诊断工具,例如血压计和听诊器,这样我产生了两三个对人类性选择的猜测。尽管我承认我发现它们的可靠性比不上趣味性,但我仍然会简要地讲一下。第一,一个关于为什么人类失去了阴茎骨的理论。人类阴茎勃起后会变得十分坚硬,以至于人们常常玩笑般地质疑里面竟然没有骨头。而事实上不少哺乳动物的阴茎里确实有着硬质的骨头,即阴茎骨,来辅助勃起。另外,这在我们的灵长类近亲中也很常见,就连我们最近的堂兄黑猩猩都有一块。尽管那是非常小的一块,而且它似乎也正朝着消失的主向进化着。似乎有这样一个趋势,即在灵长类中阴茎骨倾向于消失。我们人类,以及几种猴子,都已经完全失去了阴茎骨。所以,我们实际上失去了一个能让我们祖先很容易保持阴茎坚硬的骨头。相反,我们完全依靠一个水泵系统。尽管不大应该,但这感觉上的确是一个费力的、拐弯抹角的办法。而且,不幸的是,勃起可能失败,这直接影响了一个在野外生存的雄性的遗传成功率。最简单的一个补救方法是什么?当然非阴茎骨莫属了。因此,为什么我们不能进化出一个呢?曾经秉持“基因限制”论的生物学家不能再以“噢,需要的变种根本无法出现”这样的理由逃避问题。我们的祖先一直都有那样一块骨头,直到最近我们自己丢掉了它!为什么?
勃起在人类中完全是依靠血压。很不幸我们没有理由去相信勃起的硬度就像是医生的血压计,供雌性判断雄性的健康。但我们不只是吊在血压计这一棵树上。无论任何原因,只要勃起障碍成为某种疾病的先兆,无论是身体上还是精神上,这都可以成为使理论成立的一个版本。雌性所需要的只是一个可信赖的诊断工具。医生们不会在常规体检中做勃起测试——他们更喜欢让你伸出你的舌头。但已知勃起障碍是糖尿病及某些神经性疾病的先兆,更常见的原因是一系列心理因素——沮丧、紧张、压力大、过度疲劳、缺乏自信心等。(在自然界中,你可以试想低等级的雄性会有这种困扰。一些猴子用直立的阴茎作为一种威胁信号。)这并不是毫无道理,自然选择不断改善雌性的诊断技巧,雌性可以找出各种体现在语调上或埋藏在阴茎里的关于雄性健康的线索,以及他们处理压力的能力。但是一块骨头就能毁掉这一切!任何一个人都可以在阴茎里长出一块骨头,不需要他特别地强壮或健康。因此是雌性带来的选择压力迫使雄性失去了他们的阴茎骨,因为这样一来,就只有真正健康或强壮的雄性才能够做到真正坚硬的勃起,雌性也就能不受阻碍地做出诊断。
有一点可能会引发争论。那些施以选择压力的雌性又如何去知道她们感受到的硬度是来自骨头还是血压呢?我们从一开始就认为人类勃起可以造成有根骨头在里面的感觉,但我很怀疑雌性是否真的那么容易上当受骗。她们一样经受着选择,对她们而言,该选择不是失去某根骨头,而是增长判断力。而且别忘了,雌性能够知道那根阴茎没勃起时的样子,这个对比是如此鲜明。骨头不能收缩(尽管得承认它们可以伸缩)。可能正是阴茎这两种截然不同的状态保证了其血压式勃起广告的真实性。
现在轮到“听诊器”了。想想另外一个卧室里的梦魇:打鼾。这在现在可能只是一个小麻烦,但是在历史长河中,这可能就决定了生死。在一个寂静的夜晚,鼾声会变得非常响。它可能会把捕食者从很远的地方引到打鼾者和他的群体旁边。那么,为什么现在有那么多人打鼾呢?试想一群我们的祖先正在一个更新世的洞穴里睡觉,雄性都各自以一个不同声调打鼾,雌性则在一旁坐着,她们什么都不做,只是听着(我认为雄性更容易打鼾)。这些雄性有没有为这些雌性提供他们故意准备的、经过夸张的听诊器信息呢?鼾声精确的音质和音色是否能够成为诊断你呼吸系统健康的工具呢?我不是想说只有病了的人才会打鼾,相反,打鼾就像收音机的载频一样,无论何时都会响。这是一个为提高诊断精度调校过的工具,能反映出鼻子和咽喉的状况。雌性很明显会倾向于空荡的支气管所发出的清晰的小号般的声调,而不是充满病菌的喷嚏声。但我得承认我无法试想雌性会很适应这样的鼾声。当然,人们的直觉是那么不可靠。可能这至少能成为一个医生失眠时的研究项目吧。来思考思考吧,她可能也很想试试其他的理论哦。
请不要很严肃地对待这两个猜测。只有遵从了汉密尔顿关于雌性是如何试图挑选健康雄性的原理,它们才有可能是对的。可能最有意思的事情就是它们把汉密尔顿的寄生虫理论与阿莫兹·扎哈维的不利条件理论连上了线。如果你仔细地思考了我那阴茎假说的逻辑,就会明白雄性们因为失去了阴茎骨而变得有缺陷,而且这个缺陷还不是偶然发生的。而水压的广告之所以有效,完全是因为勃起有时会失败。达尔文主义的读者肯定注意到了“不利条件”意味着什么,并且他们可能会产生很大的怀疑。我请求他们在读完后面那个注解之前不要做出判断,那个注解将透过一个全新的视角来看待不利条件原理本身。
*50 183页:……扎哈维的……截然相反的“不利条件原理”。
在第一版里我写道:“我不相信这个理论,尽管我对它的怀疑并不像我初听到它时那么肯定了。”我很庆幸我添加了那句“尽管”,因为扎哈维的理论在现在看起来比我写下那段话时更合理了。几位受人敬仰的理论学家最近开始严肃对待该理论了。更让我担心的是,这包括了我的同行艾伦·格拉芬,他在之前被描述为“令人厌烦的永远觉得自己是对的”。他将扎哈维语言上的想法转化为数学模型并宣称模型是成立的。他并不像一些人那样对扎哈维进行着难解的滑稽模仿,而是直接将扎哈维的概念转化为了数学公式。我需要讨论一下格拉芬的模型最初的ESS版本,尽管他现在正致力于一个在某种意义上能超过那个ESS模型的全遗传学版本。但这并不意味着那个ESS模型就真的错了,而是说它是一个很好的近似模型。事实上,所有的ESS模型,包括本书中介绍的,都是指同一意义上的近似。
不利条件原理可能关系到某个个体试图判定另一个个体的某项品质这样的所有情况,但我们这里只讨论雄性对雌性放送的广告。这是为了让讨论更加清晰,因为只有这些时候那些性别代词才真的有用。格拉芬指出不利条件原理包含至少四种方式,它们是合格型不利条件(任何一个无视不利条件而存活下来的雄性一定有某方面的优势,所以雌性选择它们),展示型不利条件(雄性做出一些无谓的举动来展示它那本来隐藏着的能力),限制型不利条件(只有高质量的雄性才发育出的不利条件),最后一个格拉芬最喜欢的表达方式,他将其命名为策略选择型不利条件(雄性具有关于自身品质的私人信息,而不告诉雌性,然后用这个信息来决定是否产生一个不利条件以及这个不利条件多大比较合适)。格拉芬关于策略选择型不利条件的阐述对ESS分析有用。这里并没有对雄性所采取的广告手段有什么预先假设,即无所谓广告是代价昂贵的还是不利的。相反,他们能够自由地进化出任何一种广告手段,无论诚实与否,昂贵与否。但是格拉芬证明,就算有这样一个自由的开始,一个不利条件系统很可能趋于进化上的稳定。
格拉芬的初始假设包含了以下四点:
1. 雄性的品质是有区别的。品质并不是什么模糊或势利的概念,不像你在大学或社团中遇到的那种毫无思考能力的自满情绪(我曾经收到过一封读者来信,信中说道:“我并不希望你认为这是一封傲慢的信件,但我是贝利奥尔学院的一员。”)。对格拉芬而言,品质意味着雄性会分为好的和差的,对雌性而言,与好的结为配偶而避免差的能使它们得到遗传学上的优势。好的品质意味着诸如肌肉力量、奔跑速度、找到猎物的能力、搭建一个好的巢穴的能力。我们并没有将一个雄性最终的繁殖成功率包含入内,因为这受到雌性是否选择它的影响。在这里就引入雄性最终的繁殖成功率实际上是在回避整个问题,这是一个不确定是否能从模型里胜出的东西。
2. 雌性不能直接感知雄性的品质,只能依赖于雄性的广告。到现在为止,我们还没有做出任何关于广告是否诚实的假设。诚实是另外一个不确定是否能从模型里胜出的东西,这也是我们建立这个模型的目的。例如,一个雄性可以长出一个经过填充的肩膀,来捏造一个魁梧和强壮的假象。正是这个模型将要告诉我们这种虚假信号是否能在进化上稳定,或是自然选择会推行一套正直、诚实和可靠的广告标准。
3. 并不像雌性那样只能通过观察推断,雄性从某种意义上来说“知道”自身品质,然后它们会采用一种“广告”策略,一个根据它们品质定制的广告方案。像往常一样,“知道”并不意味着认知上的知道,而是假设雄性有能够根据其自身品质而开关的基因(而且获得这个信息的专属权并不是一个无理假设,雄性的基因往往深藏于它内部的生化环境中,远比雌性用来对它们品质产生响应的基因藏得要深)。不同的雄性会采纳不同的方案。例如,某个雄性可能会采纳“展示一个大小与我的真实品质成正比的尾巴”这样的方案,另外一个则可能采取相反的方案。自然选择可以从一群由基因控制选取方案的雄性里挑选,从而就有了一个调整方案的机会。广告的程度不一定要与真实品质成正比,事实上雄性可以选择完全相反的方案。我们需要的只是雄性应该被预设为肯定会选取某些方案来“反映”它们的真实品质,而且所选取的必须符合广告的要素——尾巴的长短,或者角的大小。这就是最终将趋于进化上的稳定的方案,也是我们想要利用这个模型找出的东西。
4. 雌性也有着相同的自由去进化它们自己的方案。对它们而言,方案应该是基于雄性广告的强度来挑选雄性(请记住它们,或更准确地说它们的基因,并没有雄性那样能观察雄性品质的特权)。例如,某个雌性可能会采纳如下方案:“完全相信雄性。”而另一个可能采纳“完全忽略雄性的广告”这样的策略,或者说另一个方案:“假设广告与实际恰恰相反。”
因此,我们的概念就是雄性的不同方案都是把品质与广告强度关联起来,雌性的方案则是把配偶选择和广告强度关联起来。对双方来说,方案都是持续变化并且由基因控制的。到现在为止,我们讨论了雄性可以选择任何将它们品质与广告关联起来的方案,雌性也可以选择任何将它们的择偶标准和雄性广告关联起来的方案。在所有这些可能的雄性与雌性方案中,我们试图寻找一对进化稳定的方案。这有点像我们通过“忠诚/薄情与放荡/羞怯”模型来寻找一个遗传稳定的雄性方案和一个遗传稳定的雌性方案。这里的稳定意味着共同的稳定,即只有其中一个方案与另一方案共存时才是稳定的。如果我们能够找到那样一对遗传稳定的方案,我们就可以研究它们,看看在遵守那样法则的雄性和雌性社会里,生活究竟是怎么样的,以及,会是一个扎哈维的不利条件世界吗?
格拉芬给自己的任务就是找到那样一对共同稳定的方案。如果我接手这个任务,我可能多半会拼命地在实验室运行计算机模拟程序。我会在计算机里放入一批采用不同方案把品质和广告挂钩的雄性样本,我也会在其中放入一批采用不同方案把择偶标准和雄性广告挂钩的雌性样本。然后我会让雄性们和雌性们在计算机里面游荡,碰到彼此,当雌性选择条件满足时结为配偶,将它们的雄性和雌性方案传给它们的儿子和女儿。当然,单个个体会依据它们遗传到“品质”而存活或者死亡。随着世代交替,每一个雄性方案和每一个雌性方案的命运变化就会表现为它们在整个群体中比例的变化。有空我就会去计算机里边看看有没有什么稳定的方案正在诞生。
这个方式从原理上说得通,但实践上却存在很大的问题。幸运的是,数学家们能够通过设立几个方程式然后求解得到相同的结果。这就是格拉芬所做的。我就不去重复他的数学原理了,也不去列出他那些进一步的更加精细的假设。相反,我会直接揭晓答案。他确实找到了一对进化稳定的方案。
所以,现在的最大问题是:格拉芬的ESS模型所构建的这种世界,是扎哈维会认同的不利及诚实的社会吗?答案是肯定的。格拉芬发现确实有个进化稳定的世界同时满足扎哈维世界的几个条件:
1. 尽管可以随意选择广告策略,雄性会选择正确反映它们真实品质的方案,无论这个方案是否会暴露它们很低的品质。换句话说,在ESS模型中雄性是诚实的。
2. 尽管可以随意选择应对雄性广告的策略,雌性最终会选择“相信雄性”。在ESS模型中,雌性对雄性有着理所应当的“信任”。
3. 广告是有代价的。也就是说,如果我们可以忽略品质和吸引力的影响,一个不做广告的雄性更容易成功(通过节省能量或者变得对捕食者不那么显眼)。广告是代价高昂的,正是由于它们有代价,所以雄性才选取了这样一个广告系统。之所以选择一个广告系统,是因为它实际上能够降低发布广告者的成功概率——在所有其他情况一样的前提下。
4. 广告的代价于越差的雄性而言越高昂。同样程度的广告带给一个孱弱雄性的风险远高于其带给一个强壮雄性的风险。低品质的雄性相对高品质的雄性来说承受着由昂贵广告带来的更大风险。
这些条件,特别是第3条,是纯粹的扎哈维式的。格拉芬展示了它们在合理情况下是进化稳定的,这看起来很有说服力。但那些影响了本书第一版的关于扎哈维的批评理由也同样有说服力,它们总结说扎哈维的观点不可能在进化中实现。我们对格拉芬的结论还不能高兴得太早,至少得等我们找出那些早前的批评究竟哪里——如果真有的话——错了。是什么假设使他们得到了一个不同的结论?部分答案似乎是他们没有让他们假想的动物有机会从诸多不同策略中做出选择。这通常意味着他们对扎哈维语言观点的阐述只是格拉芬列出来的前三种阐述——合格型不利条件、展示型不利条件、限制型不利条件——中的一种。他们都没有考虑过任何第四个版本,即策略选择型不利条件。其结果是要么他们根本无法实现不利条件原理,要么就只能在特殊的、数学抽象的情况下实现,这样对他们来说又没有了扎哈维式的矛盾感。进一步来说,不利条件原理的策略选择型阐述的关键特点在于,在ESS中,高品质的个体和低品质的个体都采取相同的策略:“诚实的广告。”此前建模的研究者都假设高品质雄性和低品质雄性使用不同的策略,因此发展出不同的广告方案。相反,格拉芬在ESS模型中假设,同样都在打广告的高品质和低品质的雄性变得不同是因为它们都使用同样的策略——它们在广告中呈现出差别是因为它们都诚实地根据信号规则来反映它们的品质。
我们始终承认信号事实上可能就是一个不利条件。我们也始终理解尽管事实上是不利条件,但极端的不利条件仍然可能得到进化,特别是作为性选择的结果。扎哈维理论中也有我们都反对的观点,就是他认为选择之所以倾向于一个信号,完全是因为它对发出信号者是一个不利因素。这也是艾伦·格拉芬现在所维护的一点。
如果格拉芬是对的——我认为他是——这对整个动物信号研究都相当重要。这甚至可能导致我们对行为的进化形势的看法来一个大转弯,也可能导致我们对本书中讨论的很多观点的看法发生大的变化。性广告只是广告的一个类型。如果扎哈维——格拉芬理论正确的话,将会完全颠覆生物学家对同性对手间的、亲代与子代间、不同物种的敌人间的关系的看法。我发现这个前景很令人担忧,这意味着我们再也不能以常识为由而排除那些几乎疯狂到极点的理论了。如果我们观察到某个动物正在做一些很傻的事,比如遇到狮子后开始用头倒立而不是远远跑开,它有可能只是在对雌性炫耀。它甚至有可能是在对狮子炫耀:“我是一个拥有如此高品质的动物,试图抓住我只会浪费你自己的时间。”(见197页)
但是,无论我认为一件事多么疯狂,自然选择总会带来一些其他的看法。当危机导致的广告效应上的增长超过危机带来的生命威胁时,一个动物甚至有可能在一群流着口水的捕食者面前表演后空翻。正是由于该姿势拥有危险性,它才更有值得炫耀的价值。当然,自然选择不可能倾向于无限的危险。当那种展示变成纯粹的有勇无谋的时候,它就会受到惩罚。一个危险或代价高昂的表演在我们看来也许很疯狂,但这真的与我们毫无关系,自然选择是唯一的审判官。
第10章 你为我搔痒,我就骑在你的头上
*51 第200页:……实际上似乎只有在社会性昆虫中才可以看到这种现象。
我们曾经这么认为,可能是忘了裸鼹鼠。裸鼹鼠是一种无毛的、近乎失明的小型啮齿动物。它们生活在肯尼亚、索马里以及埃塞俄比亚干燥区域的大型地下巢穴中。它们似乎是哺乳动物世界的“真社会性昆虫”。珍妮弗·贾维斯(Jennifer Jarvis)是开普敦大学研究裸鼹鼠人工繁殖种群的先驱,肯尼亚的罗伯特·布雷特(Robert Brett)则进一步把研究延伸到了野外观察。美国的理查德·亚历山大和保罗·舍曼(Paul Sherman)也做出了更多关于人工繁殖种群的研究。这四个学者已经承诺会联合出版一本书,我就是热切等待此书的读者中的一员。当然,这种评论是建立在阅读了仅有的几篇发表的论文以及聆听了保罗·舍曼和罗伯特·布雷特的研究讲座的基础之上。我也很荣幸地应时任哺乳动物馆馆长的布里安·伯特伦的邀请,参观过伦敦动物园裸鼹鼠种群。
裸鼹鼠生活在纵横交错的地下洞穴中。一个种群通常有70到80只个体,但有时也能有上百只。一个种群所占据的洞穴网络总长度相当于2到3英里(约为3.2到4.8千米),而且一个种群一年内会挖出3到4吨的土。打洞是一个公共行为。前线工人在最前边用牙齿挖掘,将土壤交给后面的活体传送带,那是一条拥挤、喧闹的由半打粉红色小动物组成的传送带。一段时间后,前线工人会被它后面那个工人取代。
种群里只有一只雌性生育,这种情况会持续数年时间。贾维斯采纳了社会性昆虫的术语,将它命名为女王,在我看来的确应该这样做。这个女王只和两三个雄性结合。所有其他的个体无论什么性别都是不育的,就如同昆虫里的职虫。并且,就像很多种社会性昆虫一样,如果女王被研究人员取走,一些原本不育的雌性开始变得拥有生育条件,并为女王的位置展开战斗。
那些不育的个体被称作“职虫”,当然这也是足够公平的。职虫包含了两个性别,就如同白蚁那样(但和蚂蚁、蜜蜂和黄蜂不同,在它们中职虫都是雌性的)。裸鼹鼠工人实际需要做的事取决于它们的体形。贾维斯把最小的那些称为“普通职虫”,它们负责挖掘、运送泥土、养育幼崽以及让女王安心于生育。相对于同样大小的啮齿动物,鼹鼠女王有着更多数量的幼崽,就像社会性昆虫女王一样。体形最大的不育者似乎除了吃喝睡就不干别的,中等体形的不育者则奉行中庸之道:裸鼹鼠们更像蜜蜂,有着连续的阶层,而不像很多种蚂蚁那样等级分明。
贾维斯最开始把那种体形最大的不育者称为游手好闲者。但它们真的什么都不做吗?有实验室和野外观测的证据显示它们是士兵,在有危机的时候保卫种群,蛇正是裸鼹鼠主要捕食者。它们扮演的角色也可能是像“蜜罐蚁”一样的“食物罐”(见198页)。裸鼹鼠是自食粪者,这只是用礼貌的方式描述它们吃彼此的粪便(当然不只吃这个,否则就会违背宇宙的规律)。可能那些大体形的个体的价值就在于当食物丰富时在身体内储存粪便,这样一来当食物匮乏时它们就相当于是紧急粮仓——一种粪便给养部门。
对我来说,裸鼹鼠最神奇的一点在于,尽管它们在很多方面都和社会性昆虫相似,但它们并没有像蚂蚁或白蚁里那样的带翅膀的年轻生殖者。它们当然也有生殖个体,但一开始这些生殖者并不会飞起来,把基因传播到一片新的土地上。根据现有信息,裸鼹鼠的种群扩张只会发生在原巢穴的旁边,继续扩展该地下洞穴系统。看起来似乎它们并没有像带翅膀的生殖者那样远距离传播个体。我的达尔文主义直觉对此很惊讶,这确实很让人怀疑。我的第六感告诉我某一天我们可能会发现一个由于某种原因到目前为止被忽略了的传播个体。去指望传播个体长着实体化的翅膀显然是不现实的!但它们可能会在很多方面为地上的生活而非地下生活做着准备。例如它们可能是长毛的而非裸的。裸鼹鼠并不像普通哺乳动物那样调节自己的体温,它们有点像“冷血的”爬行动物。可能它们通过社会化来调节温度——另一个与白蚁和蜜蜂的相似点。或者它们是利用任何一个好地窖都具备的恒定温度?与地底工人不同,我假想中的传播个体很可能会是通常意义上“恒温的”个体。试想,一些到现在为止还被划分为一个完全不同种类的已知多毛啮齿动物,有没有可能最终成为裸鼹鼠中尚不为人所知的哪一个阶层呢?
而且,这样的事情确实发生过,例如蝗虫。蝗虫是蚱蜢的变体,它们一般过着蚱蜢那种独居、神秘、隐居的生活。但在特定条件下,它们会发生彻底的变化——这往往也很恐怖。它们会失去自身的伪装而变得有着清晰的条纹。你几乎可以把这想象成一种警告。一旦发生改变,它们就会继续变下去,它们的行为也同样会改变。它们会放弃之前的独居生活而集结在一起,带来一种恐怖的征兆。从那传奇般的《圣经》里的蝗灾,到现代文明的今天,没有任何动物是如此可怕,以至于被当作了人类繁荣的摧毁者。那虫群数以百万计,这一收获者群体能够同时收割10英里(约19千米)宽的田地,它们有时候日行数百英里,每天能吞下2000吨的谷物,留下的只有饥荒和废墟。现在我们可以谈谈祼鼹鼠里可能存在的类似物了。一个独居个体与它的群居化身之间的区别就像是两个蚂蚁阶层之间的区别那么大。进一步来说,正如我们对祼鼹鼠中“失落的阶层”的假设,直到1921年,蚱蜢化身博士和它们的蝗虫怪人还被划分为两个不同种类的生物。
哎,但是时至今日这些哺乳动物专家看上去并不特别像会犯那样的大错。我还需要说的是在地表偶尔也能发现正常的、未变化的裸鼹鼠,可能它们旅行的距离超过了我们一般的想象。但在我们完全放弃“变形的生殖者”这一怀疑之前,蝗虫的例子也提供了另外一个可能。可能裸鼹鼠确实会产生变化的生殖者,但是要在特定条件之下——而这样的条件在最近数十年都没有发生。在非洲和中东,蝗灾仍然是很有威胁性的,正如《圣经》记录的那样。但是在北美,事情就不一样了。那儿的几种蚱蜢也能够转化为群居蝗虫。但是,可能是因为条件没达到,在20世纪北美都没有蝗灾发生(尽管另一种完全不同的虫灾,蝉造成的虫灾依旧在肆虐。令人困惑的是,美国人在口语中将蝉称作“蝗虫”)。然而,就算今天在美国发生一次真正的蝗灾也没有什么特别值得惊讶的:火山并没有熄灭,它只是休眠了而已。但如果不是我们还有世界其他地方的历史性的文字记录了这类信息的话,这就会令人恐惧和惊讶了。因为就众人所知,这些都只是一般的、独居的、无害的蚱蜢。裸鼹鼠会不会就像是美国蚱蜢一样,有能力产生出一个独立的传播阶层,但是使其产生的条件因为某种原因在这个世纪再也没出现过?19世纪的东非可能遭受着像旅鼠一样在地表迁徙的多毛鼹鼠群的灾害,但这没有为我们留下任何记录,或者可能它们已经被记录在当地部落的传说与史诗之中了?
*52 第202页:……膜翅目雌虫同它的同胞姐妹的亲缘关系比它同自己子女的更密切。
这听上去有些矛盾,汉密尔顿为膜翅目这一特例提出的“¾亲缘关系”这一难忘且精彩的假说,但这却使他那更加普遍与基础的理论陷入难堪。膜翅目¾亲缘关系刚好简单到对于任何人来说都很容易理解,但又恰好难到每个人都很高兴自己理解了这个概念,并急切地想向其他人传播。这是一个好的“觅母”。如果你不是通过阅读了解汉密尔顿,而是通过酒馆里的对话,那么你很有可能除了单倍二倍性以外什么也听不到。现在每一本生物学教科书,无论其对亲属选择的讲解是多么简要,都会使用一段来讲“¾亲缘关系”。被誉为关于大型哺乳动物的社会行为研究的世界级专家之一的同事曾经向我承认,多年以来,他都以为汉密尔顿的亲属选择就是这个¾亲缘关系假说而已!这样导致的结果是,如果一些新的事实让我们对¾亲缘关系假说的重要性产生怀疑,人们很容易认为这个证据就能推翻整个亲属选择理论。这就像是一个伟大的作曲家在谱写一部前所未有的长篇交响乐,但是在其中一个旋律的中间出现了一个破音,然后马上被一群街边小贩发现并给出一片嘘声。整部交响乐就被这一段旋律代表了。如果人们开始讨厌这段旋律,他们就认为自己讨厌整部交响乐了。
以琳达·加姆林(Linda Gamlin)最近发表在《新科学家》杂志上的一篇关于裸鼹鼠的文章为例,这篇文章从其他方面看都很有意义,但是文章的高水准为其中暗讽裸鼹鼠和白蚁以某种方式违背了汉密尔顿的假说的段落所破坏了,而这仅仅是因为它们不是单倍二倍体!很难让人相信该作者真的读过汉密尔顿的两篇经典论文,因为单倍二倍体的论述只在其50页中占了4页。她一定是通过第二手资料来了解的——我希望那不是《自私的基因》。
另外一个明显的例子和我第6章的注解中讨论的蚜虫士兵有关。正如在那里解释的那样,因为蚜虫形成同卵双胞胎的群体,所以很容易联想到它们之间会有利他主义的自我牺牲。汉密尔顿在1964年发现了这一点,但是在解释那奇怪的事实时遇到了些麻烦。以当时的了解来看,无性繁殖的动物没有任何利他主义行为的特别倾向。当发现蚜虫士兵时,这本该是一个与汉密尔顿理论完全和谐的声音。但是宣布此发现的第一批文章把它当作对汉密尔顿理论的一种挑战,蚜虫不是单倍多倍体!这是多么讽刺啊。
当我们把注意力转向白蚁时,讽刺的事情还会继续发生。白蚁也常常被视为汉密尔顿理论的瑕疵。汉密尔顿于1972年提出了一个非常睿智的理论来解释为什么它们变得具有社会性,这可以被看作单倍多倍体假说的一个聪明的类比。这个“循环近亲繁殖理论”(cyclic inbreeding theory)一般被归功于S.巴茨(S. Bartz),但他是在汉密尔顿第一次发表相关论文7年之后才发明的这个理论。按照他的一贯个性,汉密尔顿已经忘记了是他自己第一个想出的巴茨理论,而我得把他自己的文章砸在他脸上才能使他相信这一点!毋庸讳言,这个理论本身是那么有趣,以至于我都有点后悔没能在第一版中讨论它。我现在来纠正这个失误。
我说过这个理论是单倍多倍体假说的一个聪明的类比。这句话的意思是:站在社会性进化的角度来看,单倍多倍体动物的关键特征在于,从遗传学角度上来说一个个体与它的亲兄妹比其与它的后代更亲一些。这就决定了它能留在父母的巢穴内并照顾弟弟妹妹,而不是离开巢穴生育自己的后代。汉密尔顿想出了一个理论来解释为什么对白蚁来说亲兄妹间的血缘关系可能也比亲代与子代之间的关系更近。近亲繁殖为这个问题提供了线索。当动物与亲兄妹结合后,它们所产出的后代变得更加具有遗传学上的一致性。任何一个实验小白鼠品系中的小白鼠都有着与同卵双胞胎一样的相似性。这是因为它们都是经过漫长的兄妹交配过程繁育出来的。它们的基因组变得高度纯合(专业术语):它们每一个基因座上的两个基因几乎都是完全一样的,而且和其他所有该品系的个体在相同基因座上的基因都是一样的。我们很少在自然中见到长期的近亲繁殖式交配,但其中有一个重要的例外——白蚁!
一个典型的白蚁巢穴中会有一对王室夫妻:国王和女王。它们只与对方交配直到它们其中一个死去。到那时他或者她的位子就会被它们后代中的一员取代,并继续与剩下的父母之一进行近亲繁殖。如果王室夫妻双双死亡,它们就会被一对近亲兄妹夫妻取代,然后不断这样继续下去。一个成熟的群体很可能失去过很多国王与女王,因此几年以后后代很可能是极度近亲繁殖的结果,就像试验小白鼠那样。一个白蚁巢穴的平均纯合度和平均亲缘系数都会随着时间流逝而越积越高,王室生殖者也不断地被它们的后代或兄妹取代。但这仅仅是汉密尔顿论证的第一步,接下来才是精彩的部分。
任何一个社会性昆虫种群的最终产物都是一个新的带翅膀的生殖者,它会飞出父母的巢穴,交配并构建一个新的种群。当这些新的年轻国王和女王交配时,很大概率它们不会是近亲繁殖。事实上,似乎存在一个负责此事的特殊同步协定,一个区域里不同的白蚁巢穴都在同一天产生带翅膀的生殖者,似乎是为了鼓励远系繁殖。因此,考虑一下由A种群的年轻国王与B种群的年轻女王交配后的遗传学结果。双方都是高度近亲繁殖的产物,双方都相当于近亲繁殖的实验室小白鼠,但是,因为它们是经过了不同的、独立的近亲繁殖过程,它们在遗传学上是不一样的,就像是属于不同品系的近亲繁殖的小白鼠。当它们互相交配后,它们的后代会变得高度杂合,但也非常一致。杂合意味着大量基因座上的两个基因都是彼此不同的。一致性杂合就意味着几乎每一个后代都是以相同的方式杂合的。从遗传上来说,它们与它们的兄妹几乎是一样的,但同时它们仍旧是高度杂合的。
现在让我们跳过一段时间。新的种群在最初的王室夫妻的带领下茁壮成长,巢穴里充满了完全一样的杂合的年轻白蚁。想想当王室夫妻中的一个或两个死去时会发生什么。第一次近亲繁殖产出的世代将会比之前的世代更加具有多样性。无论我们考虑的是兄妹交配、父女交配还是母子交配,对所有情况来说原理是一样的,但考虑兄妹交配是最简单的。如果兄妹都是相同的杂合子,它们的后代将会有着高度的基因组合多样性。这符合最基本的孟德尔遗传学原理,这一基本原理对所有动植物都是一样的,而非仅仅对白蚁有效。当你将一致的杂合个体互相杂交或者将它们与亲代纯合体品系杂交,从遗传学上来看,就天下大乱了。其原因在任何一本基础遗传学教科书中都能找到,我就不多说什么了。从我们现在的角度来看,最重要的结果是在白蚁种群发展的这个阶段,某个个体在遗传学上与它的亲兄妹比与它的子女更亲。就像我在单倍二倍体的膜翅目中看到的一样,这正是进化出利他主义的不育职虫阶层的一个可能的先决条件。
但就算没有任何原因去期待个体们与它们的兄妹比与它们的后代更亲,也有很好的理由去期待它与两者一样亲。让这个观点成立所需的唯一条件就是要有一定程度上的一夫一妻制。在这一点上,以汉密尔顿的角度看来,最令人惊讶的是没有在更多的物种中出现照顾它们弟弟妹妹的不育工人。正如我们逐渐认识到的那样,更加广泛存在的是一种不育工人现象的稀释版本,被称为“在巢内帮忙”。在很多种鸟类和哺乳动物中,年轻个体会在离开种群组建自己的家庭之前,留在父母身边待上一两个季节,同时帮忙照看它们的弟弟妹妹。同样的基因也存在于弟弟妹妹的体内。假设受益者都是亲(而不是半亲)兄弟姐妹,从遗传上来说,给亲兄妹的每一盎司食物带来的收益和给后代相同的食物带来的收益是一样的。但这样的前提是其他条件都相同。如果我们想要解释为什么在巢内帮忙只发生于某些物种中,我们就需要审视那些不同点了。
例如,试想一种在中空树木中筑巢的鸟类。因为供应很有限,这些树十分宝贵。如果你是一只父母仍然健在的年轻小鸟,父母很可能占有着一棵这样的中空树木(至少最近它们曾经占有过一棵,要不然你就不会存在了)。因此你很可能生活在一个逐渐繁荣的中空树木中。在这个孵化场诞生的新生婴儿都是你的亲弟弟妹妹,从遗传上来说它们对你而言和你的子女一样亲。如果你独自离开,那么找到另一棵中空树木的概率很低。就算你找到了,你生育的后代们在遗传上来说也不会比你的弟弟妹妹们更亲。同样一份努力,投入到你父母巢穴的价值就会超过投入到建立一个你自己的巢穴。这些情况可能就更青睐兄妹关爱——“在巢内帮忙”。
除了所有这些,某些个体,或是每一个个体都会在某些时候,必须离开并寻找新的中空树木,或是其在该物种中的等价物。要使用第7章中“孵化以及照顾”的术语,其中一些必须负责孵化,否则就没有幼崽以供照顾了!这里要说的并不是“要不然该物种就会灭绝”,而是在一个由纯粹的照顾基因占主导地位的种群,孵化基因就会开始有优势。在社会性昆虫中,生育者的角色由女王和雄性来扮演,它们就是那些离开巢穴进入世界来寻找新的“中空树木”的青年。这也是它们为什么会有翅膀的原因,尽管在蚂蚁中工蚁并没有翅膀。这些生育阶层一生都有其特殊使命。那些在巢内帮忙的鸟类和哺乳动物则采用另外一种方式。每一个个体在生命中的某个阶段(大多是在第一个或前两个成年季)都会成为“工人”,这时它们会帮忙照看弟弟妹妹,而在其生命的其他阶段它都立志成为一名“生殖者”。
那上一个注解所描述的裸鼹鼠又怎么样呢?尽管它们的关注要点中并不包含实质的中空树木,但是它们的存在将这个关注要点或“中空树木理论”带向了极致。它们故事的关键就在于热带草原下食物供应的零散分布。它们的主食是地下的块茎。这些块茎可能会非常巨大也埋藏得非常深。这种植物的单个块茎的重量可能会超过1 000只裸鼹鼠的体重。一旦发现,就可以持续为该裸鼹鼠群提供数月甚至数年的食物。但问题就出在找寻这些块茎的过程中,因为它们随机散布在整个大草原地下。对于裸鼹鼠来说,找寻一个食物来源十分困难,但一旦找到什么都值了。罗伯特·布雷特曾经计算过,如果一只裸鼹鼠想自己去寻找那样一个块茎,它需要挖掘的距离可能会磨光它的牙。一个拥有数英里繁忙巡逻着的洞穴的大型社会性种群则是一个非常有效率的块茎矿场。每一个个体都因作为矿工团队的一员而获益。
一个由数十名工作者合作管理的大型洞穴系统就是一个类似于我们假想的“中空树木”一样的关注要点,一点也不逊色!如果你生活在这样一个繁荣的公共迷宫中,并且你的母亲仍然不断地在其中生育你的弟弟妹妹,那么事实上你想要离开并创建一个家庭的欲望就会很低。就算一些新生幼崽只是半个亲兄妹,“关注要点”理论仍然足够使年轻个体留在家中。
*53 第204页:他们发现雌雄比例令人信服地接近于3∶1的比例……
理查德·亚历山大和保罗·舍曼曾写过一篇文章批评特里弗斯和黑尔的方法与结论。他们同意在社会性昆虫中偏向雌性的性别比例很常见,但是却不同意该比例是3∶1。他们倾向于另外一个针对偏向雌性的性别比例的解释。就像特里弗斯和黑尔的解释,这个解释也是由汉密尔顿首先提出的。我发现亚历山大和舍曼的理由相当令人信服,但又不得不承认我内心觉得一个像特里弗斯和黑尔那么漂亮的工作不可能全是错的。
艾伦·格拉芬给我指出了另一个关于本书第一版对膜翅目性别比例讨论的令人担忧的问题。我在《延伸的表型》(75——76页)一书中解释了他的观点。下面是一个简单的摘要:
在任何一个可以想到的群体性别比例中,那些潜在的职虫始终在照看兄妹还是照看子女中保持中立。因此我们可以假设群体性别比例是偏向雌性的,甚至假设它符合特里弗斯和黑尔预言的3∶1的比例,因为职虫与它的姐妹相对于与它的兄弟或任一性别的后代都要更亲,那么看起来在这样一个雌性偏向的性别比例中,它应该“更愿意”照顾兄妹而不是子女:当它选择兄妹的时候,难道它没有得到最有价值的妹妹吗(外加一小撮无用的弟弟)?但是这个观点忽略了在这样一个雄性稀少的群体中雄性繁殖能力的巨大价值。职虫们可能和它的兄弟们并不那么亲,但如果雄性在整个群体中很稀少,每一个这样的兄弟都有相对更高的可能性成为未来世代的祖先。
*54 第213页:如果一个种群所处的ESS地位最终还是驱使它走上灭绝的道路,那么抱歉得很,它舍此别无他途。
卓越的哲学家,已故的J. L. 麦凯(J. L. Mackie)曾经关注过一个有趣的事实,即我的“骗子”与“傻瓜”种群可以同时达到平衡。“别无他途”可能是因为一个种群达到了一个导致其灭绝的ESS。麦凯提出了另一种观点,某些类型的ESS比起另外的类型可能更容易导致灭绝。在这个特别的例子中,骗子和傻瓜双双达到进化上的平衡:一个种群可能会在骗子的平衡点稳定或者也可能在傻瓜的平衡点稳定。麦凯的观点是在骗子平衡点稳定的种群更容易走向灭绝。因此,就有可能存在一种更高层面的,“ESS之间的”,对相互利他主义有利的选择。这可以发展成一个有利于一种类群选择理论的主张。与大部分类群选择理论不同,这个主张可能真的能行。我在《捍卫自私的基因》一文中说明了这个主张。
第11章 觅母:新的复制因子
*55 第221页:我会将赌注押在这样一条基本原则上,即一切生命都通过复制实体的差别性生存而进化的定律。
我关于宇宙中所有生命都会按照达尔文主义进化的赌局现在在我的文章《普遍的达尔文主义》(‘Universal Darwinism’)及《盲眼钟表匠》的最后一章中得到了更完善的论证。我指出所有曾经出现过的达尔文主义替代物从原理上来说就不可能解释有组织的复杂生命。这是一个普遍性的论断,并不需要基于任何我们知道的生命的细节。正因如此,它常被一些平庸到认为操作一根热的试管(或冰冷的泥靴)就是做出科学发现的唯一方法的科学家批判。一个批评者抱怨说我的主张是“哲学性”的,就好像这是一种谴责。无论哲学性与否,事实是无论他还是其他人都没有在我的文章里找出任何问题,并且像我所做的这样的“原理上的”主张,并非和现实世界没有丝毫关系。这些主张可以比那些基于特定研究的主张更加有影响力。如果这些主张是正确的,那其中的理由就告诉了我们关于宇宙任何地方的生命的一些重点信息,而实验室里的或野外的研究都只能告诉我们在这里发现的那类生命的信息。
*56 第221页:meme(觅母)
“觅母”这个词语似乎成了一个很好的觅母,它现在有着广泛的应用,并且已经在1988年入选未来版本《牛津英语词典》考虑加入的词汇列表。这使我在反复说我在人类文化方面的野心渺小到几乎不存在时更加不安。我真实的志向完全在另外一个方向上,我也承认这些志向有点大。我想要主张对于有着微小错误的自我复制实体,一旦它们诞生在宇宙任何一个角落,它们就有了几乎无限的能力。这是因为它们倾向于成为达尔文选择的基础,只要有足够的世代,达尔文选择能够逐渐地建造出非常复杂的系统。我相信在适当的条件下,自我复制因子们会自发地聚集在一起,建立能够带着它们运动以及能促进它们进一步复制的系统或者是机器。《自私的基因》前10章主要关注复制因子中的一种,即基因。我在这一章中讨论觅母是为了使复制因子更加一般化,也为了指出基因并不是这个重要类别中唯一的一员。我并不清楚人类文化环境是否真的拥有能使达尔文主义运转的要素,但通常这都是我的次要关注内容。如果读者选取这本书时觉得DNA分子并不只是满足达尔文主义进化理论的唯一实体,那么第11章就可能会获得成功。我的目的是把基因切得更小,而非重构一个关于人类文化的宏大理论。
*57 第222页:……觅母应该被看成一种有生命力的结构,这不仅仅是比喻的说法,而是有学术含义的。
DNA是硬件中自我复制的部件。每一个部件都会有一个特定的结构,该结构有别于其他竞争者。如果大脑中的觅母是基因的一个类比,它们必须是一个自我复制的脑部结构,一个能在不同大脑中重建自身的真实神经连接的形式。我总是觉得很难大声地说出这一点,因为我们对大脑的了解远逊于基因,因此我们必然对那样的大脑结构究竟是什么样感到很模糊。所以当我最近收到一份由德国康斯坦茨大学的胡安·德利厄斯(Juan Delius)所写的文章时,我感到十分地放松。与我不同,德利厄斯并不需要感到自卑,因为他是一名著名的脑科学家而我完全和脑科学不沾边。因此,我很高兴他能够勇敢地发表一个可能是觅母的神经硬件的详细图片来证明这一点。他所做的很多有趣的事情之一,就是以远比我更刨根问底的态度去探索觅母和寄生虫的类比,更准确地说,寄生虫在这里是指以有害的寄生虫为一个极端,以有益的“共生体”为另一个极端的整体。因为我自己对寄生虫基因对宿主行为的“延伸的表型”效应的兴趣(请看本书第13章以及《延伸的表型》的第12章),我很热衷于这个想法。顺便说一句,德利厄斯强调了觅母和它们的效果(“表型”)的清晰区别。他也反复强调了共适应的觅母组合体的重要性,其中觅母是根据它们的共同适应性而被选择的。
*58 第224页:《友谊天长地久》(Auld Lang Syne )
“Auld Lang Syne”不知不觉地成为我所选择的例子中一个非常好的样板。这是因为传唱中几乎普遍都出现一个错误,一个变异。特别是现在,乐曲里的叠句总是被唱为“For the sake of auld lang syne”(为了友谊地久天长),其实伯恩斯(Burns)实际填的是“For auld lang syne”(为友谊地久天长)。一个觅母化的达尔文主义者立刻就想知道那插入的短语,“the sake of”(了)究竟有什么“生存价值”。请记住我们并不是在找寻能让唱改版歌词的人们更好存活的方式,我们是在寻找能使这个变化本身在觅母库里能够更好存活的方式。每一个人都在孩提时代学会了这首歌,但他们并不是通过阅读伯恩斯的填词,而是通过在新年夜听到这首歌而学会的。在某个时间点以前,可能所有人都唱着正确的版本。“For the sake of”肯定是以罕见变异的方式出现的。我们的问题是,为什么这个一开始罕见的变异传播得如此隐蔽,以至于它最终成了觅母库里的标准?
我觉得答案不难找到。“s”的嘶嘶声非常突出。教堂唱诗班的训练就是要发“s”时越轻越好,要不然整个教堂都是嘶嘶的回音。在一座大教堂里,你往往能在听众席的最后面听到站在圣坛上的神父的轻声私语,尽管这只是零星的“s”声。“sake”中的另外一个辅音“k”也是一样具有穿透力。试想有19个人都在正确地唱着“Forauld lang syne”时,在房间某个角落,有一个人错误地唱着“For the sake of auld lang syne”。一个第一次聆听此歌的小孩很想加入合唱,但是他不清楚歌词。尽管几乎所有人都唱着“For auld lang syne”,但是“s”的嘶嘶声以及“k”的爆破声强势地占据了小孩的耳朵,当叠句再次响起时,小孩也开始唱起了“For the sake of auldlang syne”。这个变异的觅母就这样占据了另一个媒介。如果当时还有其他的孩子在场,或者是不熟悉歌词的成年人,他们很有可能在下一次叠句来临之时,也改为那个变异的版本。这并不是因为他们“偏好”那个变异的版本,他们真的是不知道歌词并且真诚渴望能够学会它。就算那些知道准确歌词的人愤愤地用他们的最高音量吼出“For auld lang syne”(就像我做的那样),但这个正确的歌词并没有十分突出的辅音,而就算别人不自信地低声吟唱变异的版本,他们的声音也能够很轻易地飘进大家的耳朵里。
一个与之相似的例子是《Rule Britannia》(统治吧,不列颠尼亚!)。该合唱正确的第二行歌词应该是“Britannia,rule the waves”(不列颠尼亚,统治这片汹涌的海洋)。尽管不是那么普遍,但它经常会被唱成“Britannia rules the waves”(不列颠尼亚统治着这片汹涌的海洋)。这里,这个觅母中冥顽不化的“s”的嘶嘶声同时被另外一种因素强化着。诗人詹姆斯·汤普森(James Thompson)用意应该是祈使式的(Britannia, go out and rule the waves!意为不列颠尼亚,走出去统治这片汹涌的海洋吧!),或者是虚拟式的(let Britannia rule the waves,意为让不列颠尼亚来统治这片汹涌的海洋),但这句话非常容易就被当作陈述句(Britannia, as a matter of fact,does rule the waves,意为不列颠尼亚,事实上统治着这片汹涌的海洋)了。这个变异的觅母因此就比其取代的原始版本多了两个不同的生存价值:听上去更突出和理解起来更容易。
这个假说必须经过实验才能算是最终测验。我们应该可以故意地以一个非常低的频率将嘶嘶的觅母插入觅母库中,然后观察它凭借自身的生存价值散布开来。要不我们几个开始唱“God saves our gracious Queen”?
*59 第224页:如果说觅母这个概念是一个科学概念,那么它的传播将取决于它在一群科学家中受到多大的欢迎。它的生存价值可以根据它在连续几年的科技刊物中出现的次数来估算。
如果这句话被理解为“吸引性”是决定是否接受一个科学观点的唯一标准的话,我就会开始厌恶这句话了,毕竟有些科学关键事实上是正确的,其他的则是错误的!它们的正确性和错误性都是能得到验证的,它们的逻辑可以用来仔细分析,它们真的不像是流行音乐、宗教或者是朋克发型。尽管有社会科学以及科学逻辑,一些差劲的科学观点依然被广泛传播,至少是在一定时期内广泛传播。有一些好的观点会在最终占领科学界之前蛰伏许多年。
我们可以在本书介绍的主要观点中找到一个这样先经过蛰伏然后快速发展的典型例子,即汉密尔顿的亲属选择理论。我觉得这是可以让我们试试通过清点期刊引用来测量觅母传播的想法的一个恰当的例子。在第一版中我指出(102页):“他在1964年发表的两篇有关社会个体生态学的论文,位列迄今为止最重要的文献之列。我一直难以理解,为什么一些个体生态学家如此粗心,竟忽略了这两篇论文(两本1970年版的有关个体生态学的主要教科书甚至没有把汉密尔顿的名字列入索引)。幸而近年来有迹象表明,人们对他的观点又重新感兴趣。”我在1976年写的这段话,让我们来追踪其后几十年里这个觅母的复兴吧。《科学引文索引》是一个非常奇特的出版物,在其中你能任选一个发表过的文献,然后通过一览表查询在特定一年里引用此文的文献数量。这是为了方便追踪某一特定主题的文献。大学任命委员会开始习惯于把其当作比较求职者科学方面成就的一个粗略和简便(过于粗略也过于简便)的方法。通过清点从1964年以来汉密尔顿文章每年的被引用次数,我们就大概能追寻到他的观点在生物学家意识里的发展(图一)。最初的蛰伏期是非常明显的,接下来在20世纪70年代,学界对亲属选择理论的兴趣有了一个显著的上升。如果真要确定一个上升的起始点的话,应该是在1973年和1974年之间。这次上升在1981年达到顶峰,之后每年的被引用次数开始在一个范围内无规律地波动。
一个觅母化的说法渐渐地成型,该说法认为对亲属选择理论的兴趣上升是因为1975年和1976年出版的一些专着导致的,而这幅图的拐点在1974年似乎使这个说法成为一个谎言。恰恰相反,这个证据可以用来支持另一个非常不同的假说,即我们总是会关注那些“正在流行”的、“它的时代已经来临了”的观点。从这个方面来看,这些70年代中期的书可能只是潮流的征兆而非是其根本原因。
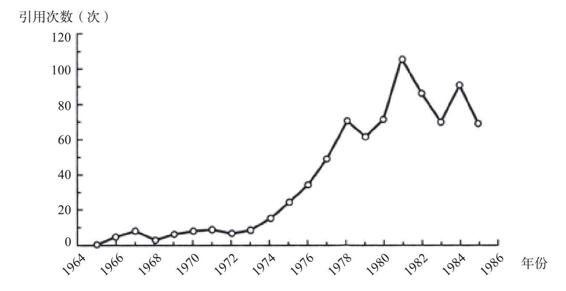
图一
很有可能我们面临的实际情况是一个在很早就开始的长期的、低起点的并呈指数级加速增长的潮流。验证这个简单的指数级假说的一个办法是使用对数刻度画出被引用次数的累计图。任何一个增长过程,如果增长速率与原先规模成正比的话,就被称为指数级增长。流行病的传播就是一个典型的指数级增长过程:每一个人呼出的病毒都会传染给其他几个人,这些人中每一个又会传染给同样多的人,因此被感染者的数量就会以一个越来越快的速率增长。指数曲线的特点就是当以对数刻度作图时,它会成为一条直线。尽管不需要以累计的方式去画出对数图像,但这样做比较常规也比较方便。如果汉密尔顿的觅母的传播真像是一个正在传播的流行病的话,累计对数图中的点都应该排列成一条直线。是这样的吗?
图二中画出的那条直线就是统计意义上最拟合所有点的直线。1966年和1967年间的那个容易看见的显著上升应该可以被忽略,因为这是这类对数图像容易积累出的一种不可靠的小数效应。因此尽管还需要忽略一些小的浮动,图中的线条还是一根很好的近似直线。如果接受了我的指数级诠释,我们所面对的就是一个单独的缓慢发展的影响力爆发。这个爆发从1967年开始到1980年结束。每一本书和论文都应被同时当作这个长期趋势的征兆及原因。
顺便说一句,尽管人们几乎不可避免地会去这么想,但真的不要认为这样的增长曲线没什么大不了的。当然,就算是被引用次数每年以一个固定数值增加,任何一个累积曲线都还是会上升的。但是在对数刻度下,它们的增速会慢慢放缓,最终消失不见。图三最上面的实心曲线就代表着如果每年的被引用次数一定的话,我们将会得到的理论曲线(被引用次数取汉密尔顿的平均值,即每年37次)。这个逐渐平缓的曲线可以直接与图二中看到的直线相比较,后者代表了一个指数级的增长。所以这是在增长的基础上还有更多的增长,而非只有一个稳定的被引用速率。
图二
图三
其次,也许这并不是不可避免的,但有些人依然会忍不住想,难道科学论文的发表速率本身不是以指数级增长的吗?这样一来引用其他文献的机会也就随之以指数级增长了。又或许科学界本身的规模就在以指数级变大。为了证明汉密尔顿觅母真的有其特殊的地方,最简单的方法就是去画一些其他文献的同类图像。图三同样也展示了另外三个作品的对数累计被引用率(这三个也对本书第一版产生过巨大影响)。这包括威廉斯1996年出版的《适应性与自然选择》、特里弗斯1971年发表的关于相互利他主义的论文以及梅纳德·史密斯和普赖斯在1973年发表的介绍ESS想法的论文。这三个作品形成的曲线在整个时间段里都明显不是指数级增长的。然而,对这些作品而言,每年被引用次数也远非一致的,在某些时间段内它们依然可能是指数级增长的。例如,那根威廉斯的曲线在对数刻度下从1970年开始就接近于一条直线了,这意味着它也进入了影响力爆发的阶段。
我低估了特定几本书对汉密尔顿觅母传播的影响。不过,对这个小小的觅母研究还有一个很明显的暗示性附注。就像“Auld Lang Syne”和“Rule Britannia”的例子一样,我们这也有一个明显的变异错误。汉密尔顿1964年发表的两篇论文的正确标题应该是“The genetical evolution of social behaviour”(《社会行为的遗传进化》)。在20世纪70年代中后期,包括《社会生物学》和《自私的基因》在内的一系列出版物都错误地将其引用成“The genetical theory of social behaviour”(《社会行为的遗传理论》)。乔恩·西格(Jon Seger)和保罗·哈维(Paul Harvey)找寻了这个变异觅母的最早的出现点,他们认为这可能是一个清晰的标记,就像是同位素标记一样,可以用来追踪科学影响。他们追溯到了E. O. 威尔逊在1975年发表的有影响力的著作《社会生物学》,他们甚至为这个可能的起源找到了一些间接的证据。
正如我非常敬仰威尔逊的力作——我希望人们更多地去阅读原作而不要去读关于它的评论——关于他的书影响了我的书这一错误看法也使我义愤填膺。是的,因为我的书也包含了那个变异的引用——那个“同位素标记”——它看起来就像是在警醒我们至少有一个觅母从威尔逊传播到了我这里。这并不是特别令人惊讶,因为《社会生物学》抵达英国时我正在完结《自私的基因》,正是那个我可能正在完善我的参考书目的时间段。威尔逊庞大的参考书目就像是一个天赐之物,省去了我很多在图书馆中的时间。但当我偶然发现了我在1970年牛津讲座中发给学生的一个陈旧的蜡纸印刷的参考书目时,我的懊悔变成了狂欢。真的,上面写着“The genetical theory of social behaviour”,比威尔逊的出版物整整早了5年。威尔逊几乎不可能看过我1970年写的参考书目。这是毋庸置疑的:威尔逊和我独立地引入了相同的变异觅母!
为什么会发生这样的巧合呢?还是那样,就像对“Auld Lang Syne”进行分析一样,并不难发现一个合理的解释。R. A.费希尔最著名的著作是The Genetical Theory of Natural Selection (《自然选择的遗传理论》),这样一个简单的名字使其在遗传生物学家的世界里扎下了根。对我们来说,很难在听到前两个单词后不自觉地去加上第三个。我怀疑威尔逊和我都是这样错的。这对所有人来说都是一个愉快的结论,因为没有人会介意承认受到过费希尔的影响。
*60 第227页:觅母存在于人的大脑中,大脑就是计算机。
我们能够很明显地预测,人工制造的电子计算机也终将成为自我复制类型的信息——觅母——的一个宿主。越来越多的计算机被连起来形成了一个错综复杂的信息分享网络。它们中很多是真正地通过电子邮件交互连接在了一起,其他的则通过它们的主人互相传递磁盘来分享信息。这是一个完美的能让自我复制程序繁荣并扩散的环境。当我在写本书第一版时,我很幼稚地认为一个不良的计算机觅母只会是发源于复制正确程序时自发性地产生的一个错误,也认为这是不大可能的。哎,那是多么天真的时代。由恶意程序员故意释放的“病毒”和“蠕虫”现在已经成了全世界计算机使用者熟悉的危害。过去几年里,据我所知我自己的硬盘就被两种不同的流行病毒感染过,我就不指出这两个病毒名称了,免得给它们龌龊幼稚的制造者们哪怕一丝龌龊幼稚的满足感。我说“龌龊”是因为我觉得他们的行为对我而言在道德上几乎和那些故意污染饮用水并为了偷笑病人而传播传染病的微生物实验室技术员没有区别。我说“幼稚”,是因为这些人心理上幼稚。设计出一种计算机病毒并没有什么聪明之处,任何一个半桶水程序员都能做到这一点,而这样的半桶水程序员在现实世界中随处可见,我自己就是其中一员。我甚至不屑于解释计算机病毒是怎么工作的,那太浅显了。
我不怎么清楚该如何战胜它们。一些非常专业的程序员很不幸不得不浪费他们的宝贵时间去写出检查病毒、杀灭病毒的程序(顺便说一句,医学疫苗接种的类似物已经很接近了,甚至已经到了注射“减活品系”病毒的程度),其危险在于引发军备竞赛,病毒防护能力道高一尺,新型病毒程序就会魔高一丈。到现在为止,大多数反病毒程序都是由利他主义者编写,也不收取任何费用。但是我预感到一个新型职业正在兴起——分化出和其他任何职业一样的一个赚钱的行当——“软件医生”。他们会背上一个装满诊断和治疗软盘的黑包随时待命。我使用了“医生”这个名字,但真正的医生是来解决自然的问题,而不是解决由人恶意制造出来的问题的。我所谓的软件医生,从另一方面来讲更像是律师,是来解决本不该存在的人造问题的。到现在为止,病毒制造者们如果真有任何可以察觉到的动机,那一定是他们感觉到了一种模糊的无政府主义。我向他们呼吁:你们真想为一个赚钱行当铺平道路吗?如果不是,停下对这些幼稚觅母的玩弄,把你们还凑合的编程才能用到一些正道上吧。
*61 第228页:盲目信仰的人什么事都干得出来。
我收到了预想中的大量信仰受害者的来信,抗议我对其提出的批评。信仰是一个如此成功的根据自己的偏好施为的洗脑者,特别是对于孩子来说,很难打破对它的信念。但究竟什么是信仰?它是大脑中让人们在毫无支持性证据的情况下相信某事物的一个状态,无论该事物究竟是什么。如果有很好的支持性证据,信仰就有些多余了,因为证据将会使我们无论如何都会相信它。正是这一点,使得那经常重复的“进化论本身是一种信仰”变得如此可笑。人们之所以相信进化论,并不是因为他们无端地想要相信,而是因为那些大家都能接触到的大量的证据。
我说了“无论(信仰相信的)事物究竟是什么”,都意味着人们对完全疯狂、武断的事物有着信仰,就像是道格拉斯·亚当斯那引人入胜的《德克·金特利的全能侦探社》(Dirk Gently's Holistic Detective Agency )中的那个电子僧侣。他是专门为实践你的信念而造的,并且很在行。有一天我们碰到了他,他坚定不移地认为世界上所有的东西都是粉红色的,而不顾所有的证据。我并不认为一个个体所信仰的事物一定是疯狂的,它们有可能是也有可能不是。关键在于没有办法去确定它们究竟是不是疯狂的,也没有办法去喜欢一个信仰胜过另外一个,因为证据被明显地隔离开了。其实真正的信仰不需要证据这一事实就已经是它最大的美德了,这就是为什么我引用了多疑的托马斯的故事,这是十二使徒中唯一值得钦佩的一员。
信仰不能移山(尽管很多代的孩子都被严肃地告知了相反的事实并且相信了它们),但是信仰却能导致人们做出那样危险的愚行,因此在我看来信仰可以被看作一种精神疾病。它能让人们如此地坚信一个事物,甚至可能极端到不需要任何进一步的论证就能为其展开杀戮或者牺牲。基思·亨森(Keith Henson)创造了“觅母者”一词来指那些“被觅母占领以至于他们自己的存活都变得不重要了的受害者……你可以从来自贝尔法斯特或贝鲁特的晚间新闻中看到很多这样的人”,信仰的威力足以使人们对所有的怜悯、宽恕及人类美好的感受免疫。如果他们真的相信殉道者死后会升入天堂,信仰甚至能使他们对恐惧免疫。多么好的一个武器啊!宗教信仰需要在武器技术年鉴中占据一席之地,与长弓、战马、坦克和氢弹站在同一条线上。
*62 第232页:在这个世界上,只有我们,我们人类,能够反抗自私的复制因子的暴政。
我结论里透出的乐观主义语调引起了那些觉得这和本书其他部分不一致的批评者的怀疑。某些例子中,批评者是一些教条主义社会生物学家,小心翼翼地保护着遗传影响的重要性。在另一些例子中,批评者来自一个近乎荒谬的相反的极端,那些左翼高级神父小心翼翼地保卫着最心爱的神学偶像。罗斯、卡明和莱文廷的《不在我们的基因里》里有一个专有的怪物叫作“还原论”,而所有最好的还原论者都被认为是“决定论者”,特别是“基因决定论者”。
大脑对于还原论者来说是一个确定的生物体,其性质就产生了我们观察到的行为以及从这些行为中领会到的思想状态或者意图……这样的观点是,或应该是,完全与威尔逊和道金斯的社会生物学原理相符的。然而,要采纳这个观点就会将这些原理引入一个两难境地。首先是争辩对于自由的人,人们觉得毫无吸引力(怨恨、教条化等)的行为是先天存在的。然后开始纠结于对犯罪行为的自由主义道德考量,因为这些行为像其他行为一样是生物上决定好的。为了避免这个问题,威尔逊和道金斯许下了一个自由的愿望,只要我们非常希望,我们就能够摆脱基因对我们的控制……这实质是回归到了难堪的笛卡尔哲学,一个二重性问题。
我认为罗斯和她的同事是在谴责我们在吃蛋糕的时候同时占有着它。要么我们必须是“基因决定论者”,要么我们就相信“自由意志”,我们不能两全其美。但是,在这里我相信我是为了威尔逊教授也是为了我自己而发声,只有在罗斯和她的同事眼中我们才是“基因决定论者”。他们没有理解的(很显然,尽管这很难确定)是我们完全有可能在相信基因对人类行为施加着统计意义上的影响的同时,也相信这样的影响是能够被改变的,比如被其他影响覆盖或逆转。基因肯定对任何一个经由自然选择而进化出的行为模式都有统计意义上的影响。罗斯和她的同事们大概都同意人类性欲是由自然选择进化而来的吧,就像很多其他经过自然选择进化的东西一样。因此他们就必须得相信存在着影响性欲的基因——就像影响其他所有东西的基因一样。然而他们显然也能够在社交上需要的情况下抑制他们的性欲吧。这和二重性有什么关系?当然没有。这个根本不存在的二重性对我提倡反叛的“反抗那些自私的基因的暴政”没有丝毫影响。我们,这里指的是我们的大脑,已经和我们的基因分开并足够独立去反抗它们了。就像已经提过的一样,我们每一次使用避孕套的时候就走出了反抗的一小步了。没有任何理由不相信我们能够进行更大的反抗。
参考书目












第14、15章参考书目






第14、15章术语表
《延伸的表型》主要是为了生物学家而写作的,他们并不需要一张术语表。但是在此我们要为这两章中出现的一些科学术语提供一个简要的解释,以便更多的读者理解书中讨论的内容。
适应(adaptation)
这个术语已经发展得多少有些远离了它在一般语言应用中的含义,即“修改”的近义。“蟋蟀的翅已经适应了(从其飞行的主要功能修改成为)歌唱的功能。”这句话含蓄地表达了蟋蟀的翅被专门设计用于歌唱,同时这样的句子也让“一种适应性”这个说法变得大概有了这样一层含义:某种生物的一种对其自身有某种“好处”的特性。何为好?为何好?为谁好?这些都是比较难回答的问题,而在《延伸的表型》中有着更为详尽的讨论。适应的例子有翅和眼。需要注意的重要一点是,适应只能从自然选择中产生。
适应性景观(adaptive landscape)
这是休厄尔·莱特(Sewall Wright 1932)发明出来的很有用处的比喻,以多维度可视化的手段将不同表型的生殖适应性比作一片景观,其中的山峰即代表了更高的适应性。一个种群的进化能够在这片景观中绘制出一条路径,它通过自然选择的作用,趋向于那些山峰(适应)。这一表述方式的一些变体也为进化生物学家和群体遗传学家所使用。
等位基因(alleles,是allelomorphs的缩写)
每一个基因都只能占据染色体上的一个特定区域,即它的基因座。在每一个给定的基因座上,种群之中都可能存在那个基因的某种替代形式,这些替代形式的基因就被称为那个基因的等位基因。《延伸的表型》强调了一点,即从某种角度来看,等位基因之间是彼此竞争的关系,因为在进化的时间尺度上,成功的等位基因相对同一基因座上的其他基因而言,将在种群中的所有染色体上获得数量上的优势。
异速生长(allometry)
指身体某一部分的尺寸与身体整体的尺寸之间的一种不成比例的关系。这一比较可以是在不同个体间进行的,也可以是在同一个体不同生命阶段之间进行的。例如,体形大的蚂蚁趋向于有着相对特别大的脑袋(在人类中则是体型小的人),其脑的生长速率不同于身体整体的生长速率。从数学上来讲,身体某一部分的尺寸与身体整体尺寸的增速相比,有着指数级别的提高。
鲍德温/沃丁顿效应(Baldwin/Waddington Effect)
最初由斯波尔丁(Spalding)于1873年提出。这是一个主要为假说性质的进化过程(也称基因同化),通过这一过程,自然选择能够创造出一种假象,好像是遗传得到了某种后天获得的特性。获得对环境刺激做出响应的某种特性,这样的遗传趋势如果被选择作用青睐,那么就会导致进化出更强的针对同一环境刺激的敏感性,并最终从对这种环境刺激的需求中解脱出来。在《对于完美化的制约》一章中,我提出我们或许能够繁育出一种自发产乳的雄性哺乳动物,方法是对连续的一代代雄性施用雌性激素,并从中选择对于雌性激素有着越来越强敏感性的雄性个体。其中激素或是其他环境因素所扮演的角色,就是让潜伏的基因变化呈现出一种开放活跃的状态。
中心法则(central dogma)
在分子生物学中,中心法则是指以核酸为模版的蛋白质合成,但两者的关系不可能对调。更普遍地讲,中心法则是指基因对于生物躯体的形式施加了影响,但是躯体的形式永远不可能被转换回基因编码——后天获得的特性不可能遗传。
染色体(chromosome)
细胞中由基因组成的链,其中除了DNA之外,通常还有由蛋白质构成的复杂支撑结构。染色体只在细胞周期中的特定一些时刻才能在光学显微镜下观察到,但是它们的数量和线性度能够仅从遗传的事实中通过统计学的推理推断出来。染色体通常存在于躯体的所有细胞之中,不过在一个细胞中只有一小部分染色体处于活跃状态。在每个二倍体细胞中通常有两个性染色体以及一些常染色体(人类有44个常染色体)。
顺反子(cistron)
基因的一种定义形式。在分子遗传学中,顺反子有着精确的定义,依赖于一种专门的实验测试的结果。宽松地讲,它被用于指称负责编码蛋白质中一条氨基酸链的一段染色体。
进化稳定策略(evolutionarily stable strategy, ESS)
注意这里出现的是evolutionarily这样一个副词,而不是evolutionary这个形容词。此处使用形容词形式是一个常见的语法错误。这个概念所指的策略在一个由此策略支配的种群中运行得很好。这个定义捕捉到了这个概念直观的本质,但多少有些不够精确。要了解数学形式的定义,请参见梅纳德·史密斯的论述(Maynard Smith 1974)。
延伸的表型(extended phenotype)
一个基因对这个世界产生的所有效应。一如既往,要理解此处基因的“效应”,就要与其等位基因相比较来看。传统意义上的表型是这个概念的一种特殊形式,其中的效应被视为限定在基因所在的动物个体身上的效应。实践中,比较方便的处理方式是把“延伸的表型”限定在一定的情况之下,即表型的效应能够以正面或负面的方式影响基因的存续机会时。
基因漂变(genetic drift)
在生物一代又一代繁育的过程中,由概率而非选择导致的基因频度的变化。
基因型(genotype)
一种生物在一个或一组特定的基因座上的基因构成。有时也会宽松地使用这个概念来代表一种表型所对应的整个基因构成。
类群选择(group selection)
一种假说性质的自然选择过程,针对多个生物群体。常常被引用以解释利他主义的进化。有时也会被人与亲属选择混淆在一起。
遗传率(heritability)
一个经常被误解的术语。它是一个统计学数值,指明了在一个种群中由基因变化引起的表型特征的可变性所占的比例,与由环境或其他因素导致的可变性比例是相对的概念。
杂合体(heterozygous)
在一个染色体基因座上有不同等位基因的情况。这个概念通常被用于一个生物个体身上,此时指称一个给定基因座上的两个不同等位基因。更宽泛地说,它可能指称等位基因对个体内或种群内所有基因座平均之后的统计学上的整体异质性。
纯合体(homozygous)
在一个染色体基因座上有一致等位基因的情况。这个概念通常被用于一个生物个体身上,此时意味着这个生物个体在一个基因座上有两个相同的等位基因。更宽泛地说,它可能指称等位基因对个体内或种群内所有基因座平均之后的统计学上的整体同质性。
广义适合度(inclusive fitness)
这个概念是比尔·汉密尔顿(Bill Hamilton 1964)从数学上提出来的,是亲属选择理论的一部分。在这个理论中,一个个体的适应性是以与其共享同样基因的近亲的适应性来估算的。
亲属选择(kin selection)
一种对于基因的选择作用,选择的标准是那些导致个体对近亲有所助益的基因,其原因在于亲属之间有很高的可能性共享了这些基因。严格来说,“亲属”包括直系子代,但不幸的是,不可否认许多生物学家使用了“亲属选择”这个说法来明确指称除了子代之外的亲属。有时也有人把亲属选择与类群选择混为一谈,但两者在逻辑上不是一回事。不过,有些物种恰好以分隔开的亲属群体形式活动,那么这两者就顺带着合二为一,成为同一样东西——“亲属类群选择”。
基因座(locus)
染色体上被一个基因(或是一组可替换的等位基因)占据的位置。例如,可能会有一个决定眼睛颜色的基因座,其上可替换的等位基因编码着绿色、棕色和红色的眼睛。这个概念通常应用于描述顺反子的层次,也可指更小或更大的染色体片段。
孟德尔遗传(Mendelian inheritance)
非混合式的遗传,借助成对的分离的遗传因子(现在被称为基因)来实现,其每一对中的两个遗传因子分别来自亲代的两个个体。其在理论上的替代概念主要是“混合式遗传”。在孟德尔遗传中,基因作用在生物躯体上的效应或许会是混合的,但是基因自身不会混合,并且会完整地传递给子孙后代。
单源的(monophyletic)
一群生物如果全都是一个共同祖先的后代,且这个共同祖先也应该被划入这个群体中,那么它们就被称为单源的。例如,鸟类可能是一个单源的群体,因为所有鸟类最晚期的共同祖先很可能也被划分为鸟类。不过,爬行动物可能就是多源的,因为所有爬行动物最晚期的共同祖先可能不会被划分为爬行动物。
米勒拟态(Müllerian mimicry)
两种作为猎物的生物物种之间的一种拟态形式,两者有着同一种捕猎者,且都有着令捕猎者反感的味道,而两者之间相互模拟着对方的警示信号,比如蝴蝶的警示色彩。这两个物种并不需要是彼此有亲缘关系的。两个物种中的个体都会从拟态中受益,因为一个捕猎者一旦品尝过其中一种猎物的难吃味道,以后就会对两个物种都避而远之。随着时间推移,更多的物种也可能来模拟同样的警示信号,无论它们是否有亲缘关系,最终就会共同形成一个“拟态环”。
突变(mutation)
基因材料中一个遗传而来的改变。在达尔文进化论中,突变被认为是随机的。这并不意味着它们不是依照一定的原理而产生的,只是说突变不存在一种导向更强的适应性的明确趋势。更强的适应性只能通过选择作用才能产生,但是它需要突变作为变化的根本性来源,才能从这些变化中做出选择。
新达尔文主义(neo-Darwinism)
一个在20世纪中叶被创造出来的术语(实际上是再创造,因为这个词已经在19世纪80年代被用于指称完全与今天概念不同的一批进化论者)。这个概念的目的是要强调(在我看来是夸大了)达尔文主义的现代综合论与孟德尔的遗传学说之间的区别。这种区别于20世纪20年代和20世纪30年代产生自达尔文自己的进化论观点之中。我认为强调“新”的必要性正在减弱,而达尔文自己的说法,即“大自然的经济性”现在看来是非常现代的。
幼态延续(neoteny)
相对于性成熟的发育过程而言,躯体的发育速度在进化过程中逐渐减缓的现象。其结果就是,执行生殖行为的生物个体相当于其古代祖先的幼年时期。有假说认为进化历史中的一些关键阶段,比如脊椎动物的起源,就是来自幼态延续。
中性突变(neutral mutation)
一个与其等位基因相比既没有选择优势也没有选择劣势的突变。理论上来讲,一个中性突变可能在几代之后被“固定下来”(也就是说,在种群中的这个基因座上占据数量优势),这也是一种进化改变的形式。对于随机性的突变固定在进化中的重要性,学界存在着合理的质疑声音,但是对于突变固定在直接产生适应性方面的重要意义则不应该存在争议——一丝一毫都不应该有。
表型(phenotype)
一种生物显露出来的某种特性,是其基因与其发育成长时所处环境合力作用的产物。一个基因可能被说成是在某方面具有表型的表达,比如说眼睛颜色方面。在《延伸的表型》这本书中,表型这个概念被延伸了,涵盖基因差异所导致的一切功能性的重要后果,包括基因所在的躯体之外的事物。
多效性(pleiotropy)
一个基因座上的改变带来明显没有联系的不同表型改变的现象。例如,某一个突变可能同时影响眼睛颜色、脚趾长度和产乳量。多效性可能是一种规律,而非特例。由于我们知道发育过程的复杂性,所以多效性是一种完全可以预计到的结果。
多源的(polyphyletic)
参见“单源的”。
复制因子(replicator)
宇宙中任何可以复制自身的实体。
存续价值(survival value)
自然选择青睐于某一特性的程度。
魏斯曼学说(Weismannism)
这个学说认为永恒不灭的种系与容纳这些种系的自然演替的肉体之间存在严格的分隔。特别是,种系能够影响躯体的形态,但是反之则不行。参见中心法则。
野生型基因(wild-type gene)
或称野生型等位基因,是某个基因座上在一个种群中最为常见的等位基因。
评论集萃
为了公益
彼得·梅达沃
发表于《旁观者》(The Spectator
),
1977年1月15日
每当在动物中发现表面上的利他主义或其他非自私的行为时,业余生物学家们,也包括越来越多的社会学家很容易被诱导,说这些行为是“为了物种的利益”才进化出来的。
以一个众所周知的谜题为例。数以千计的旅鼠以冲出悬崖掉落海里这种自杀方式来控制种群数量,很明显,它们比我们对控制种群数量的必要性有更多的认识。显然就算是那些最容易轻信的自然主义者也必须问问自己,这样的利他主义是如何成为该物种行为清单上的一项的。还得考虑到一个事实,在这个伟大的人口刑罚中,有益于这样做的遗传物质会和它们的携带者一起灰飞烟灭。然而,将这看作一个谜题,并不意味着否认遗传上自私的行为有时候可能会“表现”(就像临床医生所说)为无私或利他的行动。有利于祖母般溺爱的遗传因素和与之相反的冷漠无情相比,前者可能更容易在进化中盛行,因为慈祥的祖母们在自私地促进着存在于孙辈体内的她们自己的部分基因的存活与兴旺。
理查德·道金斯是正在崛起的一代最聪明的生物学家中的一员,他委婉且专业地揭穿了一些社会生物学关于利他主义进化的假象,但这并不意味着本书以披露为主,恰恰相反,这本书把以自然选择的遗传理论为代表的社会生物学的中心问题很有技巧地重组了一遍。另外,本书行文相当流畅,更不乏风趣与博识。与本书中出现的所有熠熠生辉的优秀生物学家一样。动物的“普遍可爱性”吸引着道金斯研究动物学。
尽管《自私的基因》的特性并不包括争论,但当道金斯在揭示某些书的虚伪之时,它就成为不可或缺的一环,这些书包括了洛伦茨的《论进犯行为》、阿德里的《社会契约》以及艾贝尔——艾伯费尔德(Eibl-Eibesfeldt)的《爱与恨》(Love and Hate )。“这些书的问题是它们的作者大错特错了……因为他们误解了进化的工作原理。他们做出了一个错误的假设,即进化中重要的是要对整个物种(或整个群体)有利,而不是对每个个体(或基因)有利。”
学童们的格言本上一打诸如“一只母鸡是一只蛋制造另一只蛋的方式”之类的警句,确实是有道理的。理查德·道金斯这样写道:
本书的论点是,我们以及其他一切动物都是各自的基因所创造的机器……我将要论证,成功基因的一个突出特性就是其无情的自私性。这种基因的自私性通常会导致个体行为的自私性。然而我们也会看到,基因为了更有效地达到其自私的目的,在某些特殊情况下,也会滋长一种有限的利他主义。上句中,“特殊”和“有限”是两个重要的词。尽管我们可能觉得这种情况难以置信,但对整个物种来说,普遍的爱和普遍的利益在进化论上简直是毫无意义的概念。
我们可能会哀叹这些真相,道金斯说道,但这并不能减少它们哪怕一丁点儿的真实性。然而,我们对遗传过程中的自私性了解得越清楚时,我们就越有资格去教导慷慨、合作以及其他所有为公益奉献的美德。道金斯也更加清楚地阐述了文化上或“外源性”进化对人类的特殊重要性。
在第11章也是最重要的一章里,道金斯对自己提出了一个挑战,去构建一个对所有进化系统都能够成立的基本原理——这甚至可能是由硅原子取代了碳原子的生物体,或者是像人类一样,很多进化都是由非遗传方式而实现的生物体。该原理就是,进化是通过复制实体的净繁殖优势而实现的。对于一般的生物体,在一般的情况下,该实体就是DNA分子中被称为“基因”的片段。对道金斯来说,文化传播的基本单位就是被他称作“觅母”的东西。在第11章中,他阐述了觅母的达尔文主义理论究竟是什么。
我想为道金斯这本令人兴奋的好书添加一个脚注:奥地利生理学家埃瓦尔德·赫林(Ewald Hering)在1870年首先提出,拥有记忆功能是所有生物的基本特征。他将他的基本单位称作“觅聂米”(mneme)——精心选择的一个忠于其语源的词汇。很自然,理查德·西蒙对这个主题的阐述(1921年)完全是非达尔文主义的,所以除了成为一个历史插曲,现在它什么也不是。赫林的一个观点被与他对立的一个自然哲学家霍尔丹揶揄道:该观点就是,必有一个化合物拥有那些属性,我们现在知道它就是脱氧核糖核酸,即DNA所拥有的。
©《旁观者》,1977
自然的博弈
汉密尔顿发表于《科学》,
1977年5月13日(节选)
所有人都应该来阅读这本书,此书也的确适合所有人群。它使用绝妙的技巧描述了进化论的新面孔。最近有很多书籍,致力于以一种轻松、无碍的形式向公众传授新的生物学知识,但有时候那些知识却是错误的。在我看来,与那些书相比,本书更加严谨也更加成功。它成功地完成了那个看似不可能完成的任务,即使用简单、非技术性的英语来表达一些更深奥的、更数学的关于近期进化思潮的主题。以这些主题带来的宽广视角纵览这本书,它甚至能使很多自认为已经清楚的研究型生物学家感到惊讶和振奋。至少,他们会对这个评论者感到惊讶。再重复一次,这本书对任何人都是很容易阅读的,哪怕他没有任何科学背景。
就算不是故意去假冒内行,但当某人阅读一本领域与自己研究兴趣相似的畅销书时,他几乎都会情不自禁地去找碴:这个例子用得不好,那个观点太模糊了,这个概念是错的,很多年前就被弃用了。但本书令我无话可说。这并不是说这里面没有问题——这对于一个从某种意义上来说是现炒现卖的作品是不大现实的——而是指其中的生物学内容从整体上来看是朝向正确的方向的,其可能存在问题的论点也是非教条主义的。该作者对自己观点的谦逊表达缓和了很多批评,读者们总能在这里或那里找到这样一个让他们很开心的提议:如果他们不习惯给出的模型,他们应该能找出另一个更好的模型。能在畅销书中做出如此严肃的邀请生动地反映了该话题的新颖性。不可思议的是,那些还未得以验证的简单观点可能可以迅速地解决一些进化论里的古老谜题,而这样的可能性真的存在。
那么,进化论的新面孔究竟是什么呢?从某种程度上来说,这就像是对莎士比亚的一个新的阐述:尽在书中,却又视而不见。然而,我需要补充的是,出现问题的那些新观点在达尔文的进化论书卷中并没有它们在自然界中隐藏的那么深,我们更像是有个20年的时间差而不是100年。例如,道金斯从那些我们了解得比较清楚的变化多端的螺旋状分子起笔,但达尔文甚至不了解染色体或是染色体在生物繁殖过程中演出的奇异舞蹈。但就算是20年,这时间也已经长的足够令人惊讶了。
第1章概括地描写了这本书想要解释的现象,并展示了它们在哲学上以及实际上对人类生命的重要性。一些有趣的或惊恐的动物事例抓住了我们的注意力。第2章回到了原始汤中,聚焦第一群复制因子。我们看着它们增殖并变得日益精巧。它们开始为物质竞争,彼此之间战斗,甚至裂解或者吃掉对方。它们将自己以及它们的收获及武器都藏在一个防御围栏中。使用这些围栏并不只是使它们免受竞争者和捕食者的困扰,也是保护它们免受它们即将侵入的恶劣环境的影响,这样它们继续移动、定居,变成奇怪的形态,被泼洒在沙滩上、土地中,以及沙漠和无尽的风雪里。在这些长久以来生命不能存在的边界之中,原始汤被一遍遍的泼洒了数百万次,形成了一个异常奇特的模型多样性。终于,它们被泼洒成了蚂蚁和大象,山魈与人类。第2章总结道,考虑到这些古老复制者结合而成的一些终极后代:“保存它们正是我们存在的终极理由……今天,我们称它们为基因,而我们就是它们的生存机器。”
很有说服力并令人激动,读者可能会这样想,但是这真的非常新颖吗?好吧,到现在为止可能还不是,但是进化过程并没有停滞于我们的身体。更重要的仍然是那些能使生物在这拥挤的世界中存活下来的技能,这个世界有着超乎想象的微妙,微妙到远超生物学家们在陈旧的、过时的、为了物种利益而适应的模型中所能展望到的。大致来说,这本书剩余部分的主题正是这种微妙性。举一个简单例子,鸟类的歌声。这看上去似乎是一个非常没有效率的安排:一个天真的唯物主义者在研究鸫属中的一个种,探寻它们是如何活过严酷的冬天、食物匮乏时期等问题时,他会觉得发现它们中的雄性会发出艳丽的歌声就如同降神会中真的降下灵魂那样不现实。(更进一步的思考会使他觉得,这个物种竟然存在雄性这一个事实同样是不现实的,而这实际上是这本书另一个主要话题:就像鸟类的歌声一样,过去我们对性的功能的论证太简单了。)然而在任何一个鸟类种群中,一整队的复制者都在提醒自己去为这样的表演做好准备。道金斯在某处引述了一个更加不同寻常的歌声,座头鲸的歌声能传遍整个海洋。但对这个歌声,我们的所知远比我们对鸫属歌声的了解还要少,它们是干什么的,是向谁发出的?目前掌握的证据表明,它可能是对鲸类联合反抗人类的一首赞美诗——如果它真是,可能也对鲸有好处吧。当然,正是这些复制者队伍中的另外一队现在举办起了交响音乐会,这些声音当然有时也会穿越大洋——从空间上的身体位置来看的话。而这些身体本身又是由一些更加复杂的团队根据计划组装并运行的。如果道金斯是对的,魔法师用镜子做的事和自然所做的相比简直不值一提,除了冻结着的原始汤,自然根本没有任何有希望的初始原料。这本书将协助塑造一个新的面孔,这正是在这本书以及其他一些近期出版的书籍(例如威尔逊的《社会生物学》)中想要表达的生物学面孔。它闪耀着一种希望,这些生命中很遥远的细节可能很快就会被更完善地组合在一起,进而成为一个更加普遍的形式,这个形式会包含最简单的细胞壁,最简单得多细胞机体以及乌鸫的歌声。如果不是细节上的组合,这就将是一个基本要素的结合(有信仰的人和非马克思主义者可以将这句话颠倒过来,如果这样做更适合他们的话)。
然而,有一种印象需要避免,即认为这本书是门外汉或穷人的《社会生物学》。首先,它有很多原创概念,其次,通过着重强调威尔逊没有提及的社会行为的博弈论方面,它从某种意义上来说平衡了威尔逊的巨著。“博弈论”并不是一个非常准确的词,特别是在低层次的社会进化情况下,因为基因本身并不据理行动。然而,我们已经很清楚,在所有层面上博弈论与社会进化的概念结构都有着重要的相似性。这种互相间取长补短意味着这个领域非常新颖并且仍在发展中:例如,直到最近我才知道博弈论已经将一个概念命名为“纳什均衡”,这个概念大概可以对应于“进化稳定策略。”道金斯正确地处理了进化上的平衡性,这也同时成为他很多关于社会生物学新观点的重要基础。在任何社会环境下,社会行为中博弈型要素以及社会适应性都来自个体策略的成功,而一个个体策略的成功是建立在它能击败与该个体相互作用者所采取的策略的基础上。无论周遭情况如何,都要在一个给定的环境中攫取最大利益,这种对适应性的追寻会带来一些非常令人惊讶的结果。例如,与大部分动物的情况相反,如果必定有一方去做这件事的话,鱼类往往是由雄性去守卫卵和孩子。谁能想到这样一个重要问题可能取决于一个非常小的细节,即哪一个性别首先在水里释放它的配子。然而道金斯和他的同事利用特里弗斯的概念,解释了为什么那样一个时间上的细节,甚至可能就是几秒钟,就对整个现象是如此关键。再举一个例子,我们难道不会认为,因为有着雄性的照顾,在一夫一妻制下的鸟类中的雌性,会比多配偶制下的雌性产下更大的蛋吗?但事实恰恰相反。道金斯在他那有点危言耸听的关于“两性战争”的章节里,再一次运用了稳定性的概念解释了利用(在这个例子中是雄性所为),并一下就使得这个怪异的关系变得自然起来。他这个观点,就和他很多其他观点一样,还没有得到证实,因此还可能有其他的,更加重要的原因。但是他给出的这个观点,从他全新的角度来看是非常明显的,所以这个观点值得我们注意。
在博弈论的教科书中你找不到任何的博弈,就如同你在现代几何中很难看到圆圈和三角一样,粗略一看都是些代数:博弈论从一开始就是一门技术型学科。因此,暂且不谈内在的细节,能够像这本书一样,在包含了如此多的博弈论情形之时不用到公式,这本身就是一个文学上的奇迹。那些博弈论的情形甚至包括了其外在感受与质量,更不用说内在细节了。费希尔在介绍他那本关于遗传的伟大著作时写道:“我没有任何一个努力是为了使这本书更加易读。”在那本书中,瓢泼大雨般的公式以及深奥且非常简洁的句子很快就把人们打懵了,使他们陷入了沉默。阅读完《自私的基因》,我现在觉得费希尔可以做得更好,尽管必须得承认,那样他就得去写另外一类的书了。看上去似乎那些经典群体遗传学的构成要件都能在一篇散文中变得比之前更加有趣。(其实,霍尔丹在这方面做得比费希尔是要好一些,但他的书也没那么深奥。)但真正引人注目的是这个新的、更加社会学的研究生命要素的方法能绕过多少令人生厌的数学,这些数学主要是由在怀特、费希尔和霍尔丹带领下的群体遗传学主流群体带来的。当我发现道金斯分享同我一样的对费希尔的评论时,我感到非常惊讶,他把费希尔誉为“20世纪最伟大的生物学家”(我认为这是一个非常罕见的评论),但我同样也惊讶于发现他几乎没怎么反复讲费希尔的书。
最后,在第11章中,道金斯开始了关于文化进化这一非常迷人的主题。他创造了“觅母”一词(meme,“mimeme”的缩写)作为文化上“基因”的等价物。就像很难去界定这个词的范围一样——这显然会比“基因”更难,尽管那已经够糟了——我推测这个词很快会成为生物学家的常用词,并且希望它也会被哲学家、语言学家以及其他专家运用。它可能会像“基因”一样融入日常对话之中。
Excerpted with permission from W. D. Hamilton,SCIENCE 196:757-59(1977)© 1977 AAAS
基因和觅母
约翰·梅纳德·史密斯写于《伦敦图书评论》,
1982年2月4—18日。(节选自对《延伸的表型》的评论)
《自私的基因》不平凡的一点在于,尽管是按畅销书的类型来写的,却仍然为生物学做出了原创性贡献。进一步说,它本身的贡献是另类的。不像戴维·拉克的经典著作《罗宾的生活》(The Life of the Robin )——同样是一本通俗形式的原创性贡献——《自私的基因》并没有提出任何新的事实。它也没有包含任何新的数学模型,实际上它根本就没有数学。它提供的是一个新的世界观。
尽管大家已经广泛阅读并欣赏了这本书,但它还是引起了深深的敌意。我相信,很多这样的敌意都是因为误解而来,或者更准确地说是因为多个误解才导致的。其中,最根本的就是没能理解这本书是要说什么。这是一本关于进化过程的书,它不关乎道德,或者政治,或者人类科学。如果你并不关心进化是怎么来的,并且不能理解为什么有的人会如此严肃地去关心一些和人类无关的事,那就不要读它:它只会导致你不必要的愤怒。
然而,假设你对进化很感兴趣,理解道金斯这样做的原因的最好办法就是去了解20世纪60年代到70年代进化生物学家们进行辩论的要点。这包含了两个相关主题,“类群选择”和“亲属选择”。“类群选择”的辩论开端于温-爱德华兹,他认为行为适应是通过“类群选择”进化而来——也就是说,通过一些群体的生存和其他群体的灭绝……
在几乎相同时间,汉密尔顿提出了另外一个关于自然选择是如何实现的问题。他指出,如果基因导致了它的携带者牺牲自己的生命去拯救其他几个亲属的生命,比起没有做这样的牺牲,会有更多的该基因的拷贝制品会存在下去……为了定量地模拟这个过程,汉密尔顿引入了“广义适合度”这个概念……这不仅仅包含了一个个体自己的后代,而且还包含了另外的接受该个体帮助的亲属的后代,这会根据关系的远近亲疏而相应变化……
道金斯在承认我们欠汉密尔顿债的同时,也认为他错误地做出了保留拟合度这一概念的最后尝试,他觉得汉密尔顿应该更加聪明地去采用一个纯粹基因的眼光看待进化。他催促我们去注意“复制因子”与“载体”之间的根本区别。复制因子是指其精确结构在每次繁殖的时候得到复制的实体,载体则是暂时性的,并且不能被复制,但是载体的性质可以被复制因子影响。我们所熟悉的主要复制因子就是核酸分子,通常是DNA分子,它们是基因和染色体的组成部分。常见的载体则是狗、果蝇及人的身体。因此,假使我们观察到一个结构,例如眼睛,它是为了适应观察而出现的,我们就可能很有理由去问眼睛究竟是为了谁的好处而进化出来的。道金斯认为,唯一合理的答案是它是为了复制因子的更好发展而进化出来的。尽管像我一样,他非常偏向于以个体优势而非群体优势来作为一个解释,但他更应该偏向于以复制因子的优势至上。
©约翰·梅纳德·史密斯,1982