來自理查德·道金斯與王巖合著的《祖先的故事》(2006,第二版)。感謝王巖提供圖片。
科學家與政客不同,能夠以錯為樂。政客如果改變了主張,會被人說成是“反覆無常”。託尼·布萊爾(Tony Blair)就誇口說自己“從未開過倒車”。科學家一般來說也願意看到自己的觀點被證明是正確的,但是偶爾一兩次的錯判同樣能夠為自己贏得尊重,尤其是當他們能夠優雅地承認自己的錯誤時。我從未聽說過有哪位科學家被指責為反覆無常。
在某種意義上,我很願意找到一種方式來收回《自私的基因》一書的核心思想。在基因組學的世界中已經發生了如此之多激動人心的事情,那麼一本出版於40年前的以基因為題的書如果還不至於被徹底摒棄的話,必然需要接受大幅度的修改——這似乎是不可避免的事情,甚至很是誘人。然而在這本書中,“基因”的定義比較特殊,它是為進化量身定做的,而不以描述發育問題為目的——若非如此,這本書就真要大改特改了。本書中基因的定義來自種群進化生物學家喬治·C. 威廉斯 [1] ,他已然仙逝,但無疑是本書的英雄。同樣離我們而去的還有約翰·梅納德·史密斯 [2] 和比爾·漢密爾頓。威廉斯認為:“基因是染色體材料上任何一個可能存在得足夠長久的代際,並且是可以當作自然選擇的單位的部分。”我從這個定義中得出了一個多少有些好笑的結論:“嚴格來講,這本書的書名應該是……《染色體有點自私的一大部分以及更為自私的一小部分》。”胚胎學家關心的是基因會如何影響表型,我們新達爾文主義者的關注點則是實體在種群中的頻度發生的改變。這些實體在威廉斯看來就是基因(威廉斯後來稱之為“抄本”)。基因是可以計數的,而其出現頻度是其成功與否的一種測度。本書的一個核心思想是:生物個體不具備上述討論的基礎。單個生物體的基因頻度都是100%,因而無法“當作自然選擇的單位”。同樣,複製單元也無法扮演這樣的角色。如果非要說生物個體是自然選擇的單位,這其中的意味也很不一樣,實際上是把生物個體視為基因的“載具”。生物個體成功與否的測度是其所攜帶的基因在未來代際中出現的頻度,而其奮力去爭取最大化的那個量值是漢密爾頓所說的“廣義適合度”(inclusive fitness)。
一個基因要獲得這些數值層面的成功,就需要在生物個體身上表現出表型效應的價值來。一個成功的基因會在很長一段時期內表現在許多個體身上,它能夠幫助這些個體在環境中存活得足夠長久,令繁育下一代成為可能。不過,這裡的環境指的並不僅僅是身體之外的外部環境,諸如樹木、水體、天敵等等,其實還包括了內部環境,特別是其他基因——自私的基因與其他基因共享了一系列生物個體的身體,不僅遍佈種群,而且跨越代際。由此,必然的結果是,自然選擇會青睞那些在有性繁殖種群中其他基因的陪伴下共同繁榮的基因。從本書的主張來講,基因的確是“自私的”。但基因同時也能夠與其他基因合作,它們所共享的不僅僅只是某一個特定的生物體的身體,還是這個物種的基因庫所產生出來的一般意義上的所有身體。一個有性繁殖種群是由相互兼容、彼此合作的基因所組成的聯合體——它們今天會合作是由於許多世代以來在先祖類似身體中的合作已經讓它們得以繁榮。需要了解的重點在於(這一點常常被人誤解),合作性之所以得到青睞,並非是因為一組基因天然地作為一個整體接受選擇,而是因為單個基因是單獨接受選擇的,但這個選擇過程的背景是該基因在身體內有可能接觸到的其他基因,也就是說物種的基因庫內的其他基因。一個有性繁殖物種的每一個個體都是從基因庫中抽取自己的基因的。在一系列個體的身體裡,這一物種(而非其他物種)的這些基因會不斷地彼此相遇,彼此合作。
我們仍不完全清楚究竟是什麼原因推動了有性繁殖的起源。但是有性繁殖的一個結果就是,物種可以被創新性地定義為:相互兼容的基因所組成的合作聯合體的棲息地。正如在《基因的延伸》這一章所指出的:合作的關鍵在於,對於每一代而言,一個身體裡所有基因共享的那個去往未來的出路如同“瓶頸”一般——那是它們渴望搭乘著前往下一代的精子或卵子。“合作的基因”也是一個同樣恰當的書名,而且如此一來,這本書就根本不用做任何修改了,我估計人們因誤解提出的許多質疑也可以因此而避免了。
另一個不錯的書名是“不朽的基因”。“不朽”比“自私”更有詩意,同時也抓住了本書的一個關鍵性問題。DNA(脫氧核糖核酸)複製的高保真度對於自然選擇而言是一個基本要素,也就是說突變是罕見的。高保真度意味著,基因作為準確的信息拷貝能夠存續數百萬年之久,成功的基因如此,不成功的基因則不行——而這正是成功性的定義。不過,如果一段遺傳信息可能的生命週期很短的話,兩者的差異就不會很顯著。換個角度來看,每一個活著的生物個體從其胚胎發育期開始就是由一些基因建造出來的,而這些基因能夠追溯到許多許多代際以前的許多許多祖先個體身上。活著的生物繼承了這些曾經幫助過許多許多祖先存活下來的基因,這就是為什麼活著的生物具備存活下去所需的一切,並且能夠繁衍下去。它們所需的東西具體是什麼,是因物種不同而各異的——捕獵者或是獵物,寄生蟲或是宿主,水生或是陸生,棲息在地下或是森林的樹冠層——但是普遍性的原則仍是相同的。
本書的一個核心論點是由我的朋友,偉大的比爾·漢密爾頓提出並完善的。我至今仍在為他的離世感到哀痛。我們認為動物應該不僅僅要照料自己的孩子,而且還要照料其他有血緣關係的親屬。對此,有一個我很喜歡的簡潔的表述方式,就是“漢密爾頓法則”:如果一個利他性的基因能夠在種群中擴散開來,那麼必然滿足以下條件,即利他者的成本C要小於受益者獲得的價值B與兩者之間的親緣度係數r之積。這裡的r是一個介於0與1之間的比例。對於同卵雙胞胎來說,r的取值是1,子女或同胞兄弟姐妹則是0.5,孫子孫女、異父或異母的兄弟姐妹,以及同胞兄弟姐妹的孩子是0.25,同胞兄弟姐妹生出來的堂或表兄弟姐妹之間則是0.125。但什麼情況下r是0呢?在這種定義之下,0的含義是什麼?要解釋清楚這個問題會更困難一些,但的確是很重要的,而且在《自私的基因》第一版中沒有完全給予闡明。0並不意味著兩個個體沒有共同的基因。所有人類之間共享著超過99%的基因,人與老鼠也有超過90%相同的基因,與魚有四分之三相同的基因。這些相當高的比例令許多人對於親屬選擇產生了誤解,其中甚至還包括一些傑出的科學家。但是上面這些數字並非是r的含義。比如說我與我兄弟之間的r值是0.5,那麼我與種群背景之中一個可能會與我競爭的任意成員之間的r值就是0。為了對利他主義的進化進行理論化的分析,堂兄弟姐妹之間的r值為0.125只是在與種群參照背景(r = 0)相比較的情況下選取的。這裡的參照背景是指種群中其餘的個體,利他主義可能也曾在他們身上體現過:雖然面對食物和空間時是競爭對手,但面對物種的生存環境時是歷經久遠歷史的夥伴。這些數值,0.5或是0.125等,指代的是在親緣關係趨近於零的種群參照背景之上的額外的親緣關係。
威廉斯所定義的基因是一種可以計數的東西,你可以隨著一個物種的代際更替去計算它的數量變化,這無關其分子層面上的本質是什麼。比如說,基因的本質是被基本呈惰性的“內含子”(會被翻譯機器忽略 [3] )分隔的一系列“外顯子”(被表達的部分),然而這一事實並不影響威廉斯所定義的基因。分子基因組學是一個迷人的領域,但是它並未對進化的“基因視角”產生巨大的衝擊,而這種視角正是這本書的中心主題。為了說明這一點,我們可以換個說法:《自私的基因》的觀點很可能對於其他星球上的生命也是能夠成立的,即便那些星球上的基因與DNA毫無關係。不過,現代分子遺傳學的具體內容,以及關於DNA的具體研究成果,是可以有辦法納入基因視角之中的。結果你會發現,它們實際上證明了我關於生命的觀點的正確性,而非對其產生懷疑。對於這一點,我會稍後再繼續探討。而現在,我要換個跨度有點大的話題,它始於一個非常具體的問題,而這個問題顯然能夠代表一大票類似的疑問。
你與伊麗莎白二世女王(Queen Elizabeth II)之間的親緣關係有多近?對我來說,我知道自己是她的十五重堂兄弟姐妹的孫輩。我們共同的祖先是第三代約克公爵理查·金雀花(Richard Plantagenet,1412—1460)。理查的兒子之一是英王愛德華四世(King Edward IV),伊麗莎白女王是他的後代。理查還有一個兒子是克拉倫斯公爵喬治(George,傳說是在一桶烈性白葡萄甜酒中淹死的),我是他的後代。很多西方人可能不知道自己與女王的親緣關係其實比十五重還近,我亦如此,門口的郵遞員也是如此。有很多種方法可以讓我們成為某個人的遠房表親,或者讓我們都成為彼此的親戚。我知道自己是妻子的十二重堂兄弟姐妹的孫輩(共同祖先是喬治·黑斯廷,第一代亨廷頓伯爵,1488—1544),但是很有可能我們還能通過某種未知的不同方式成為血緣關係更近的親戚(從各自祖先查下來的不同路徑),而且絕對還有許多其他的方式讓我們成為血緣關係更遠的親戚。我們所有人都是如此。你和女王可能既是九重堂兄弟姐妹的六世孫輩,又是二十重堂兄弟姐妹的玄孫輩,還是三十重堂兄弟姐妹的八世孫輩。我們所有人,無論生活在世界上的哪個地方,不僅僅是彼此的遠房親戚,而且還有幾百種連接親緣關係的路徑。這只不過是用另一種方式表達:我們所有人都是種群的背景,我們之間的親緣關係指數r趨近於零。我可以依照有據可查的一條路徑去計算自己與女王之間的r值,但是根據r值的定義,最終算出來的數值會非常接近於零,因而完全起不了什麼作用。
導致所有這些令人頭昏腦漲的多重親緣關係的原因在於性。我們有兩位父母,四位祖父母,八位曾祖父母,越往上越多,近乎天文數字。如果你不斷乘以2,一直計算到征服者威廉的年代,你的(以及我的、女王的、郵遞員的)祖先數量將至少是個十億數量級的數字,比當時全世界的人口數量還多。這個計算本身就證明,無論你是從哪裡來的,我們都共同擁有許多祖先(如果回溯到足夠久遠的過去,我們的祖先都是完全相同的),所以我們彼此也是很多不同形式的親戚。
如果你不是從生物個體的視角(生物學家的傳統視角),而是從基因的視角(這一整本書都在以不同的方式來提倡這種視角)來看待親緣關係這件事情的話,所有這些複雜性就都不存在了。不要再問我與我的妻子(或郵遞員,或女王)有什麼樣的親緣關係,相反,要從單獨一個基因的視角來提問,比如我的藍眼基因:我的藍眼基因與郵遞員的藍眼基因有什麼關係?像ABO血型系統這樣的多態性可以回溯到久遠的過去,甚至連猿類和猴子也有同樣的系統。站在人類A型血的基因的立場上看,黑猩猩身上與之對應的基因是其近親,而人類的B型血基因反而親緣關係更遠。在Y染色體上的SRY基因決定著動物個體是否是雄性。我的SRY基因會把一隻袋鼠的SRY基因“看作”近親。
或者,我們還可以從線粒體的視角來看待親緣關係的問題。線粒體是充斥在我們細胞之中的小東西,攸關生死。它們是無性繁殖的,保留了自己殘存的基因組(它們與自由生活的細菌有極遠的親緣關係)。根據威廉斯的定義,一個線粒體的基因組可以被看作一個單一的“基因”。我們只能從母親那裡獲得線粒體。所以,如果我們想知道自己的線粒體與女王的線粒體有怎樣的親緣關係,那麼只會有一個答案。我們可能不知道那個答案是什麼,但是我們的確知道她的線粒體與你的線粒體之間成為親戚的方式只有一種,而非從整個身體的視角來看時的幾百種不同方式。你可以一代一代回溯你的祖先,但是隻能沿著母親這條線回溯,你會得到一條單一的(線粒體的)細線,而非不斷分支的“整體生物家譜”。你還可以用同樣的方法回溯女王的祖先,追蹤她的那條母系細線。遲早,這兩條線會彼此相遇,此時只要數數每條線上過了多少代,你就能輕易計算出你與女王的線粒體親緣關係了。
你能在線粒體上做的操作,理論上也可以在任何一個特定的基因上重複,這就顯示出了基因視角與生物體視角的不同。從整個生物個體的視角來看,你有兩位父母,四位祖父母,八位曾祖父母,等等。但是,每個基因就像線粒體一樣,只有一位父母輩,一位祖父母輩,一位曾祖父母輩,等等。我有一個藍眼基因,女王則有兩個。理論上,我們能夠回溯基因的代際,找到我的藍眼基因與女王的每一個藍眼基因之間的親緣關係。我們兩個基因的共同祖先被稱為“聚結點”。聚結分析已經成為遺傳學的一個繁盛分支,而且非常吸引人。你看出來了嗎?這與整本書都在宣揚的“基因視角”是多麼相得益彰啊!我們已經不是在談論利他主義了。基因視角在其他領域也能顯示出自己的力量,比如在尋找祖先這件事情上。
你甚至可以去分析一個生物個體的身體裡兩個等位基因之間的聚結點。查爾斯王子也有藍色的眼睛,我們可以假定他的15號染色體上有一對不同的藍眼等位基因。查爾斯王子的兩個藍眼基因分別來自他的父母,這兩個基因之間有多近的親緣關係呢?對此,我們知道一個可能的答案,而這僅僅是因為王室的家譜比我們大多數人的家譜記錄得更為詳盡。維多利亞女王也有藍眼,而查爾斯王子可以有兩種方式算作維多利亞女王的後代:從他母親那邊通過國王愛德華七世來算,或從他父親那邊通過黑森的愛麗絲公主來算。有50%的可能性,維多利亞的一個藍眼基因剝離出了兩個拷貝,一個給了她的兒子愛德華七世,另一個給了她的女兒愛麗絲公主。這兩個子代基因更進一步的拷貝很容易一代代傳到伊麗莎白二世女王和菲利普親王身上,從而在查爾斯王子身上重新組合在一起。這就意味著,查爾斯兩個基因的“聚結點”是維多利亞女王。我們不知道,也不可能知道,查爾斯的兩個藍眼基因是否真是這樣的情況。但是從統計學上來說,他的很多對等位基因肯定都會回溯到維多利亞女王身上聚結。同樣的推理也適用於你的一對對基因和我的一對對基因。即便我們可能沒有查爾斯王子那樣清晰記錄下來的族譜可供查詢,但是你的任何一對等位基因理論上也能找到它們的共同祖先,那就是它們從一個單一先祖基因上“剝離”開的聚結點。
下面要說的是一件有意思的事情。雖然我無法找到自己任何一對特定等位基因的確切聚結點,但是理論上來說,遺傳學家們能夠對任何一個個體身上的所有對等位基因做聚結點的分析,考慮歷史上所有可能的親緣路徑(實際上不是所有的可能路徑,因為實在太多了,所以只能是抽取一個統計上的樣本),從而得出這個個體整個基因組的聚結模式。位於劍橋的桑格研究所的李恆(Heng Li)和理查德·德賓(Richard Durbin)發現了一件非常棒的事情:一個單一個體的基因組中所有成對等位基因的聚結模式就可以給我們足夠多的信息,讓我們得以重建其整個物種在之前歷史中的種群統計學細節,確定那些重要事件的發生時間。
在我們關於成對基因聚結的討論中,兩個等位基因中的一個來自父親,一個來自母親。這裡“基因”的含義比分子生物學家所說的基因更為靈活一些。實際上,你可以說聚結遺傳學家迴歸到了一個概念上,有點類似於我之前所說的“染色體有點自私的一大部分以及更為自私的一小部分”。聚結分析研究的對象是一塊塊的DNA序列,它可能比分子生物學家所理解的單個基因更大,也可能甚至更小,但是它們仍舊能被視為彼此的親戚,是從一個有限數量的世代之前的共同祖先那裡“剝離”出來的。
當一個(上述意義上的)基因“剝離”成為自己的兩個拷貝,並分別傳給兩個子女時,這兩個拷貝的後代可能會隨著時間的流逝而積累不同的突變。這些突變可能是“不會被注意到的”,因為它們不會表現為表型上的區別。兩者之間突變的差異與分離之後經歷的時間是成正比的。這一事實,也就是所謂的“分子時鐘”,已經被生物學家們很好地利用起來了,可以測量巨大的時間跨度。此外,我們去計算親緣關係的這些成對基因不一定要有著相同的表型效應。我有一個來自父親的藍眼基因,與之成對的等位基因則是來自母親的棕眼基因。雖然這些基因不同,但它們必定在過去的某個時候有一個聚結點:在那個時刻,我父母的共同祖先的一個特定基因剝離成了兩個拷貝,分別傳給了這位祖先的兩個孩子。這次聚結(不同於維多利亞女王藍眼基因的兩個拷貝)是很久很久以前的事情,從而讓這對基因有足夠長久的時間來積累差異,尤其是導致不同眼睛顏色的差異。
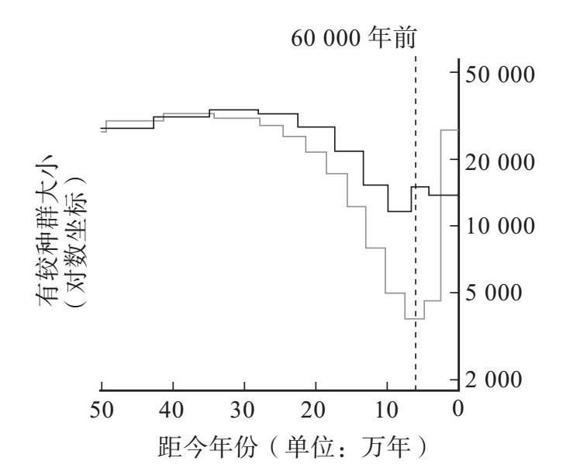
我剛才提到過,一個生物個體的基因組聚結模式就能夠用以重建之前歷史中的種群統計學細節。實際上,任何一個個體都可以用來做這件事。碰巧我的整個基因組已經為了錄製《性、死亡與生命的意義》這個電視節目進行了測序,這期節目曾於2012年在四頻道上播放過。王巖(Yan Wong)是我寫作《祖先的故事》(The Ancestor’s Tale )的共同作者,我對於聚結理論的所知以及許多其他方面的知識都是從他那裡學到的。王巖利用這個機會對我的基因組做了李/德賓式計算,而且只用了我自己的基因組而已,就推導出了人類的歷史。他發現大量的聚結點出現在大概6萬年以前。這表明我的祖先所在的那個交配種群在6萬年前是一小群人。當時的人口少,那麼一對現代基因聚結到當時某位祖先身上的概率就高。較少一些聚結點出現在30萬年前,說明當時的有效種群要更大一些。這些數字可以繪成一條曲線,縱軸是有效種群的大小,橫軸是時間。下圖就是他發現的模式,它與該技術的創始人在任何一個歐洲人的基因組上可以得到的模式都是相同的。
來自理查德·道金斯與王巖合著的《祖先的故事》(2006,第二版)。感謝王巖提供圖片。
圖中黑線顯示了估算出來的不同歷史時期的有效種群的大小,依據的是我的基因組數據(我父親與我母親之間基因的聚結)。它表明,我的祖先的有效種群大小曾經在大約6萬年前顯著減小。灰線顯示了分析一個尼日利亞人的基因組得到的模式。它顯示出差不多同一時期的種群也有所縮小,但程度沒有那麼劇烈。或許無論是什麼災禍導致了這一下降,它在非洲的下降程度都要弱於在歐亞大陸的下降程度。
順帶一提,王巖曾是我在牛津大學新學院指導的本科生,但後來我從他身上學到的東西多過了他從我身上學到的。本科畢業後,他成為艾倫·格拉芬 [4] 的研究生,而格拉芬也曾是我教過的本科生,後來成為我的研究生,而現在我稱他為我知識上的導師。所以,王巖既是我的徒子,又是我的徒孫——這恰恰從文化傳承的角度類比了我先前描述的情況,即大家能夠以不同的方式成為親戚。當然了,文化傳承的方向要比上述這個簡單的構想所代表的情況複雜得多。
總結一下,以基因的視角來看待生命是這本書的一箇中心主題,它所闡明的不僅僅是此前版本中詳細予以論述的利他和自私的進化,它還闡明瞭更為久遠而深刻的過去,那是我在最初寫作《自私的基因》時沒能覺察到的。《祖先的故事》第二版中的相關段落(主要是由我的共同作者王巖撰寫)為這個問題提供了更為全面的詳盡論述。基因視角是如此的強大,以至於一個單一個體的基因組就足以進行詳細的定量推算,得出種群的歷史性統計數據。它還有什麼其他的強大功能嗎?正如與尼日利亞人的比較所預示的,未來對於世界不同地域的人所做的此類分析,能夠為這種來自過去的種群統計學信號賦予一個地理維度。
基因視角還可能以其他方式穿透歷史的迷霧嗎?我在幾本書中曾經提出了一個想法,我稱之為“死亡的基因之書”。一個物種的基因庫就是相互支持的基因的聯合體,它們曾經一起在過去的特定環境中存活了下來,有久遠的過去,也有近世的牽絆,這就使得它們成為那些環境的一種相反印記。如果一個遺傳學傢俱備足夠的知識,就應該能夠從一種動物的基因組中讀出該物種曾經生活的這些環境的特徵。理論上來講,歐洲鼴鼠(Talpa europaea )的DNA應該能夠生動地展示出地下世界的面貌,那是一個位於地表之下的潮溼、黑暗的環境,空氣中滿是蠕蟲、腐葉、甲蟲幼蟲的氣味。如果我們知道如何去閱讀的話,單峰駱駝(Camelus dromedarius )的DNA將會展現出用編碼描繪的古老沙漠,那裡有著沙塵暴、沙丘,以及生命對水的渴望。常見的寬吻海豚(Tursiops truncatus )的DNA表達著“深海水域”“快速追逐魚類”“躲避虎鯨”等信息,不過是以一種我們在未來的某一天可能予以解讀的語言寫就的。但是同樣的海豚DNA中也包含了一些描述其更早時期所處世界的段落,相應的基因也存續了下來:那是它的祖先們在陸地上生活的時期,要小心躲避暴龍和異特龍,活得足夠久,久到能夠生育才行。在那之前,肯定還有一部分DNA描述了更為古老的生存技能,又一次帶我們回到了海洋中,那是它的先祖們還是魚的時期,被鯊魚甚至是廣翅鱟(巨大的海蠍子)追逐的時代。對於“死亡的基因之書”的研究工作應該會在未來某個時間成為活躍的熱點。它會不會成為《自私的基因》50週年紀念版的後記中的精彩內容呢?
[1] 指喬治·克里斯托弗·威廉斯(George Christopher Williams),美國進化生物學家。——譯者注(下文如無特殊說明,均為譯者注)
[2] 約翰·梅納德·史密斯(John Maynard Smith),英國著名進化生物學家、遺傳學家。
[3] 原文如此。但實際上,內含子與外顯子一起被轉錄成為信使RNA之後,還要經過細胞質內的剪切體的加工,去除不表達的內含子對應區段,才能獲得成熟的信使RNA,進入核糖體進行表達。核糖體這個“翻譯機器”在此過程中並不會“忽略”內含子。
[4] 艾倫·格拉芬(Alan Grafen),英國動物行為學家和進化生物學家。